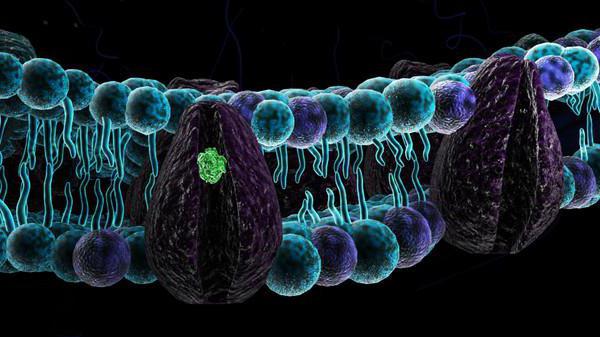
Какие выделяют три типа мембранных белков. Интегральные и периферические мембранные белки. Закрепление с помощью мембранного «якоря»
Будущее медицины - персонифицированные методы избирательного воздействия на отдельные системы клетки, которые ответственны за развитие и течение конкретного заболевания. Основным классом терапевтических мишеней при этом являются мембранные белки клетки как структуры, ответственные за обеспечение непосредственной передачи сигналов в клетку. Уже сегодня почти половина лекарств воздействуют именно на клеточные мембраны, и дальше их будет только больше. Знакомству с биологической ролью мембранных белков посвящена данная статья.
Структура и функции клеточной мембраны
Из школьного курса многие помнят устройство структурной единицы организма - клетки. Особое место в устройстве живой клетки играет плазмалемма (мембрана), которая отделяет внутриклеточное пространство от окружающей ее среды. Таким образом, главная ее функция - создание барьера между клеточным содержимым и внеклеточным пространством. Но это не единственная функция плазмолеммы. Среди других функций мембраны, связанных в первую очередь с мембранными белками, выделяют:
- Защитную (связывание антигенов и предупреждение их проникновения в клетку).
- Транспортную (обеспечение обмена веществ между клеткой и средой).
- Сигнальную (встроенные рецепторные белковые комплексы обеспечивают раздражимость клетки и ее ответ на различные воздействия извне).
- Энергетическую - преобразование разных форм энергий: механической (жгутики и реснички), электрической (нервный импульс) и химической (синтез молекул аденозинтрифосфорной кислоты).
- Контактную (обеспечение связи между клетками при помощи десмосом и плазмодесм, а также складок и выростов плазмолеммы).

Строение мембран
Мембрана клетки - это двойной слой липидов. Бислой образуется благодаря наличию в молекуле липидов двух частей с разными свойствами - гидрофильного и гидрофобного участка. Наружный слой мембран образован полярными «головками» с гидрофильными свойствами, а гидрофобные «хвосты» липидов обращены внутрь бислоя. Кроме липидов, в структуру мембран входят белки. В 1972 году американские микробиологи С.Д. Сингер (S. Jonathan Singer) и Г.Л. Николсон (Garth L. Nicolson) предложили жидкостно-мозаичную модель строения мембраны, согласно которой, мембранные белки «плавают» в бислое липидов. Эта модель была дополнена немецким биологом Каем Зимонсом (1997) в части образования определенных, более плотных участков с ассоциированными белками (липидных рафтов), которые свободно дрейфуют в бислое мембраны.
Пространственная структура мембранных белков
В различных клетках соотношение липидов и белков различно (от 25 до 75% белков в пересчете на сухую массу), и расположены они неравномерно. По расположению белки могут быть:
- Интегральными (трансмембранными) - встроенными в мембрану. При этом они пронизывают мембрану, иногда неоднократно. Их внеклеточные участки часто несут цепи олигосахаридов, формируя гликопротеиновые кластеры.
- Периферическими - расположены преимущественно на внутренней стороне мембран. Связь с липидами мембраны обеспечивается за счет водородных обратимых связей.
- Заякоренными - преимущественно расположены с наружной стороны клетки и «якорем», удерживающим их на поверхности, является молекула липида, погруженная в бислой.

Функционал и обязанности
Биологическая роль мембранных белков многообразна и зависит от их структуры и расположения. Среди них выделяют рецепторные белки, канальные (ионные и порины), транспортеры, моторы и структурные белковые кластеры. Все виды мембранных белков-рецепторов в ответ на какое-либо воздействие меняют свою пространственную структуру и формируют ответ клетки. Например, рецептор инсулина регулирует поступление глюкозы внутрь клетки, а родопсин в чувствительных клетках органа зрения запускает каскад реакций, что приводят к возникновению нервного импульса. Роль мембранных белков-каналов заключается в транспорте ионов и поддержании разницы их между внутренней и внешней средой. Например, натрий-калиевые насосы обеспечивают обмен соответствующих ионов и веществ. Порины - сквозные белки - участвуют в переносе молекул воды, транспортеры - в переносе некоторых веществ против градиента концентраций. У бактерий и простейших движение жгутиков обеспечивают молекулярные белковые моторы. Структурные мембранные белки поддерживают саму мембрану и обеспечивают взаимодействие других белков плазмолеммы.

Белки для мембраны, мембрана для белков
Мембрана - это динамическая и очень активная среда, а не инертная матрица для белков, которые в ней расположены и работают. Она существенно влияет на работу мембранных белков, а липидные рафты, перемещаясь, формируют новые ассоциативные связи белковых молекул. Многие белки просто не работают без партнеров, и межмолекулярное их взаимодействие обеспечивается характером липидного слоя мембран, структурная организация которого, в свою очередь, зависит от структурных белков. Нарушения в этом тонком механизме взаимодействия и взаимозависимости приводят к нарушению функций мембранных белков и целому ряду заболеваний, таких как диабет и злокачественные опухоли.
Структурная организация
Современные представления о структуре и строении мембранных белков основаны на том, что в мембранной периферической части большинство из них состоит редко из одной, чаще из нескольких ассоциированных олигомеризующихся альфа-спиралей. Причем именно такая структура является залогом выполнения функции. Однако именно классификация белков по типам структур может принести еще немало сюрпризов. Более чем из ста описанных белков наиболее изученным по типу олигомеризации мембранным белком является гликофорин А (белок эритроцитов). Для трансмембранных белков ситуация выглядит сложнее - описан лишь один белок (фотосинтетический реакционный центр бактерий - бактериородопсин). Учитывая высокую молекулярную массу мембранных белков (10-240 тысяч дальтон), у молекулярных биологов широкое поле для исследований.
Сигнальные системы клетки
Среди всех белков плазмолеммы особое место принадлежит рецепторным белкам. Именно они регулируют, какие сигналы поступят в клетку, а какие нет. У всех многоклеточных и некоторых бактерий передача информации осуществляется посредством специальных молекул (сигнальных). Среди этих сигнальных агентов выделяют специально секретируемые клетками), небелковые образования и отдельные ионы. Последние могут выделяться при повреждении соседних клеток и запускать каскад реакций в виде болевого синдрома, главного защитного механизма организма.
Мишени для фармакологии
Именно мембранные белки являются главными мишенями применения фармакологии, так как именно они и есть те точки, через которые идет большинство сигналов. «Нацелить» лекарственный препарат, обеспечить его высокую селективность - вот главная задача при создании фармакологического средства. Избирательное воздействие только на конкретный тип или даже подтип рецептора - это влияние только на один тип клеток организма. Такое селективное воздействие может, например, отличить опухолевые клетки от нормальных.

Лекарства будущего
Свойства и особенности мембранных белков уже сегодня используются в создании лекарств нового поколения. Эти технологии основаны на создании модульных фармакологических структур из нескольких молекул или наночастиц, «сшитых» друг с другом. «Нацеливающая» часть узнает на мембране клетки определенные рецепторные белки (например, связанные с развитием онкологических заболеваний). К этой части добавляется разрушающий мембрану агент или блокатор процессов производства белков в клетке. Развивающийся апоптоз (программа собственной гибели) или другой механизм каскада внутриклеточных превращений приводит к желаемому результату воздействия фармакологического средства. В результате мы имеем лекарство с минимумом побочных эффектов. Первые такие лекарства по борьбе с раком уже проходят клинические испытания и вскоре станут залогом высокоэффективной терапии.

Структурная геномика
Современная наука о белковых молекулах все интенсивнее переходит на информационные технологии. Экстенсивный путь исследований - изучить и описать все, что можно, сохранить данные в компьютерных базах и потом искать пути применения данных знаний - такова цель современных молекулярных биологов. Всего лишь пятнадцать лет назад стартовал глобальный проект «геном человека», и мы уже имеем секвенированную карту генов человека. Второй проект, цель которого - определить пространственное строение всех «ключевых белков», - структурная геномика - пока далек от завершения. Пространственная структура определена пока только для 60 тысяч более чем из пяти миллионов белков человека. И пусть пока ученые вырастили лишь светящихся поросят и холодоустойчивые помидоры с геном лосося, технологии структурной геномики остаются этапом научного познания, практическое приложение которого не заставит долго себя ждать.
ЛЕКЦИЯ
ТЕМА:” Введение в гистологию. Плазматическая мембрана, строение и функции. Структуры, формируемые плазматической мембраной”
Гистология в дословном переводе - это наука о тканях, однако это понятие не вмещает того действительно большого обьема материала, который освещает эта понастоящему медицинская дисциплина. Курс гистологии начинается с изучения цитологии не столько на светооптическом, сколько на молекулярном уровне, который в современной медицине логически вошел в этиологию и патогенез целого ряда заболеваний. Гистология – это и отдельные разделы из курса эмбриологии, не всей конечно, а той ее части, которая затрагивает вопрос закладки и дифференцировки тканевых зачатков. И,наконец, гистология – это большой раздел частной гистологии, то есть, раздел, изучающий строение и функции различных органов. Перечисленные разделы курса гистологии не оставляют сомнения в том, что изучение нашей дисциплины следует проводить в аспекте сохранения единства клеточного, тканевого, органного и системного уровней организации
Мы начнем гистологию с изучения эукариотической клетки, являющейся самой простой системой, наделенной жизнью. При исследовании клетки в световом микроскопе мы получаем информацию о ее размере, форме, и эта информация связана с наличием у клеток ограниченных мембраной границ. С развитием электронной микроскопии (ЭМ) наши представления о мембране, как о четко ограниченной линии раздела между клеткой и окружающей средой изменились, ибо оказалось,что на поверхности клетки имеется сложная структура, состоящая из следующих 3-х компонентов:
1. Надмембранный компонент (гликокаликс) (5-100 нм)
2. Плазматическая мембрана (8-10 нм)
3. Подмембранный компонент (зона вариации белков цитоскелета)
При этом 1 и 3 компоненты вариабельны и зависят от типа клеток, наиболее статичным представляется строение плазматической мембраны, которую мы и рассмотрим.
Изучение плазмолеммы в условиях ЭМ привело к заключению об однотипности ее структурной организации, при которой она имеет вид триламинарной линии, где внутренний и наружный слои электронноплотные, а расположенный между ними – более широкий слой представляется электроннопрозрачным. Такой тип структурной организации мембраны свидетельствует об ее химической гетерогенности. Не касаясь дискуссии по этому вопросу, оговорим, что плазмолемма состоит из трех типов веществ: липидов, белков и углеводов.
Липиды , входящие в состав мембран, обладают амфифильными свойствами за счет присутствия в их составе как гидрофильных, так и гидрофобных групп.
Амфипатический характер липидов мембраны способствует образованию липидного бислоя. При этом в фосфолипидах мембраны выделяют два домена: а) фосфатная – голова молекулы, химические свойства этого домена определяют его растворимость в воде и его называют гидрофильным
б) ацильные цепи, представляющие собой этерифицированные жирные кислоты – это гидрофобный домен.
Типы мембранных липидов. 1. Основным классом липидов биологических мембран являются фосфо(глицериды) (фосфолипиды), они формируют каркас
биологической мембраны (рис. 1).
Биомембраны – это двойной слой амфифильных липидов (липидный бислой). В водной среде такие амфифильные молекулы самопроизвольно образуют бислой, в котором гидрофобные части молекул ориентированы друг к другу, а гидрофильные к воде (рис. 2).
В состав мембран входят липиды следующих типов:
1. Фосфолипиды
2.Сфинголипиды “головки” + 2 гидрофобных “хвоста”
3.Гликолипиды
Холестерин (ХЛ) – находится в мембране в основном в срединной зоне бислоя, он амфифилен и гидрофобный (за исключением одной гидроксигруппы). Липидный состав влияет на свойства мембран: отношение белок/липиды близок 1:1, однако миелиновые оболочки обогащены липидами, а внутренние мембраны – белками.
Способы упаковки амфифильных липидов : 1. Бислои (липидная мембрана), 2.Липосомы - это пузырек с двумя слоями липидов, при этом как внутренняя, так и наружная поверхности являются полярны. 3. Мицеллы – третий вариант организации амфифильных липидов – пузырек, стенка которого образована одним слоем липидов, при этом их гидрофобные концы обращены к центру мицеллы и их внутренняя среда является не водной, агидрофобной.
Наиболее распространенной формой упаковки молекул липидов является образование ими плоского бислоя мембран. Липосомы и мицеллы – это скорые транспортные формы, обеспечивающие перенос веществ в клетку и из нее. В медицине липосомы используют для переноса водорастворимых, а мицеллы – для переноса жирорастворимых веществ.
Белки мембраны:
1. Интегральные (включены в липидные слои)
2. Периферические
Интегральные (трансмембранные белки):
1. Монотопные – (например, гликофорин. Они пересекают мембрану 1 раз), и являются рецепторами, при этом их наружный – внеклеточный домен – относится к распознающей части молекулы.
2. Политопные – многократно пронизывают мембрану – это тоже рецепторные белки, но они активизируют путь передачи сигнала внутрь клетки.
Мембранные белки, связанные с липидами.
4. Мембранные белки, связанные с углеводами.
Периферические белки – не погружены в липидный бислой и не соединены с ним ковалентно. Они удерживаются за счет ионных взаимодействий. Периферические белки ассоциированы с интегральными белками в мембране за счет взаимодействия - белок-белковые взаимодействия.
Пример этих белков:
1. Спектрин , который расположен на внутренней поверхности клетки
2. Фибронектин, локализован на наружной поверхности мембраны
Белки – обычно составляют до 50% массы мембраны. При этом
интегральные белки выполняют следующие функции:
а) белки ионных каналов
б) рецепторные белки
2. Периферические мембранные белки (фибриллярные, глобулярные) выполняют функции:
а) наружные (рецепторные и адгезионные белки)
б) внутренние – белки цитоскелета (спектрин, анкирин), белки системы вторых посредников.
Ионные каналы – это сформированные интегральными белками каналы, они формируют небольшую пору, через которую по электрохимическому градиенту проходят ионы. Наиболее известные каналы – это каналы для Nа, К, Са 2 , Сl.
Существуют и водные каналы – это аквопорины (эритроциты, почка, глаз).
Надмембранный компонент – гликокаликс, толщина 50 нм. Это углеводные участки гликопротеинов и гликолипидов, обеспечивающие отрицательный заряд. Под ЭМ – это рыхлый слой умеренной плотности, покрывающий наружную поверхность плазмолеммы. В состав гликокаликса помимо углеводных компонентов входят периферические мембранные белки (полуинтегральные). Функциональные участки их находятся в надмембранной зоне- это иммуноглобулины (рис. 4) .
Функция гликокаликса: 1. Играют рольрецепторов .
2. Межклеточное узнавание .
3. Межклеточные взаимодействия (адгезивные взаимодействия).
4. Рецепторы гистосовместимости.
5. Зона адсорбции ферментов (пристеночное пищеварение).
6. Рецепторы гормонов .
Подмембранный компонент или самая наружная зона цитоплазмы, обычно обладает относительной жесткостью и эта зона особенно богата филаментами (d 5-10 нм). Предполагают, что интегральные белки, входящие в состав клеточной мембраны, прямо или косвенно связаны с актиновыми филаментами, лежащими в подмембранной зоне. При этом экспериментально доказано, что при агрегации интегральных белков, находящийся в этой зоне актин и миозин также агрегируют, что указывает на участие актиновых филамент в регуцляции формы клетки.
Структуры, формируемые плазмолеммой
Контуры клетки, даже на светооптическом уровне, не представляются ровными и гладкими, а электронная микроскопия позволила обнаружить и описать в клетке различные структуры, которые отражают характер ее функциональной специализации. Различают следующие структуры:
1. Микроворсинки – выпячивание цитоплазмы, покрытые плазмолеммой. Цитоскелет микроворсинки сформирован пучком актиновых микрофиламент, которые вплетаются в терминальную сеть апикальной части клеток (рис. 5). Единичные микроворсинки на светооптическом уровне не видны. При наличии значительного их числа (до 2000-3000) в апикальной части клетки уже при световой микроскопии различают “ щеточную каемку”.
2. Реснички – располагаются в апикальной зоне клетки и имеют две части (рис. 6) : а) наружную - аксонему
Б) внутреннюю – безальное тельце
Аксонема состоит из комплекса микротрубочек (9 + 1 пары) и связанных с ними белков. Микротрубочки образованы белком тубулином, а ручки – белком динеином – эти белки в совокупности формируют тубулин-динеиновый хемомеханический преобразователь.
Базальное тельце состоит из 9 триплетов микротрубочек, расположенных у основания реснички и служит матрицей при организации аксонемы.
3. Базальный лабиринт – это глубокие инвагинации базальной плазмолеммы с лежащими между ними митохондриями. Это механизм активного всасывания воды, а так же ионов против градиента концентрации.
1. Транспорт низкомолекулярных соединений осуществляется тремя способами:
1. Простая диффузия
2. Облегченная диффузия
3. Активный транспорт
Простая диффузия – низкомолекулярные гидрофобные органические соединения (жирные кислоты, мочевина) и нейтральные молекулы (Н О, СО, О). С увеличением разности концентраций между отсеками, разделенными мембраной, растет и скорость диффузии.
Облегченная диффузия – вещество идет через мембрану также по направлению градиента концентрации, но с помощью транспортного белка – транслоказы. Это интегральные белки, обладающие специфичностью в отношении переносимых веществ. Это, например, анионные каналы (эритроцит), К - каналы (плазмолемма возбужденных клеток) и Са - каналы (саркоплазматический ретикулум). Транслоказа для Н О – это аквапорин.
Механизм действия транслоказы:
1. Наличие открытого гидрофильного канала для веществ определенного размера и заряда.
2. Канал открывается только при связывании специфического лиганда.
3. Канала нет как такового, а сама молекула транслоказы, связав лиганд, поворачивается в плоскости мембраны на 180 .
Активный транспорт – это транспорт с помощью такого же транспортного белка (транслоказы), но против градиента концентрации. Это перемещение требует затрат энергии.
В отличие от липидов, мембранные белки трудно классифицировать по структуре, целесообразнее подразделять основные вили мембранных белков исходя из их функций. Как правило, именно белки ответственны за функциональную активность мембран. К таким белкам относятся разнообразные ферменты, транспортные белки, рецепторы, канальные белки, белки, образующие водные поры (аквапорины), а также различные структурные и регуляторные белки, которые обеспечивают многообразные функции клеточных мембран. По биологической роли мембранные белки можно разделить на четыре группы:
1) белки-ферменты, обладающие каталитической активностью;
2) рецепторные белки, специфически связывающие те или иные вещества;
3) структурные белки;
4) белки, обеспечивающие межклеточные взаимодействия.
Белки-ферменты наиболее распространены среди всех мембранных белков. В их число входят как интегральные (мембранные АТФазы, выполняющие транспортную функцию), так и периферические (ацетилхолинэстераза, кислая и щелочная фосфатазы, РНКаза) белки. Транспортные белки создают устойчивые потоки определенных веществ и ионов через мембраны. Транспорт ионов приводит к возникновению трансмембранного потенциала во всех клетках, а так же к его изменениям в нервной и мышечной клетках. Последнее явление лежит в основе таких важнейших свойств этих тканей, как возбудимость и проводимость.
Рецепторными белками называют белки, специфически связывающие те или иные лиганды, участвующие в передаче сигналов от одних клеток к другим.Такая передача осуществляется различными способами. Часто рецепторы входят в состав более сложных мембранных комплексов, содержащих белки-исполнители. Например, в нервных и нервно-мышечных синапсах сигнальной молекулой (медиатором) является определенное низкомолекулярное вещество, а плазматическая мембрана содержит специальные рецепторные белки, соединенные с ионными каналами, изменяющие свои свойства при связывании рецептора с лигандами. Эта реакция обеспечивает проницаемость мембраны для различных ионов (натрия, калия, кальции, хлора) и формирует возбуждающий потенциал. Некоторые рецепторы (например, никотиновый холинорецептор) сами являются ионными каналами (за счет включения в рецепторный ансамбль дополнительных белковых субъединиц).
В ряде случаев рецепторный белок не является ионным каналом, но связан с внутриклеточным сигнальным каскадом, активация которого происходит при связывании с рецептором лиганда, несущего информацию. В результате активации таких рецепторов (их называют метаботропными, в противоположность ионотропным, регулирующим ионные потоки через мембрану) возникает каскад химических реакций, управляющих клеточными функциями через изменения метаболизма (отсюда и название этих рецепторов). Активация метаботропных рецепторов лигандами (их можно считать первичными сигнальными молекулами, или первичными мессенджерами) приводит к выработке в цитоплазме активируемой клетки вторичных сигнальных молекул (вторичных мессенджеров).
Структурные белки придают клеткам и органеллам определенную форму; обеспечивают те или иные механические свойства (например, эластичность) плазматической мембране; осуществляют связь мембраны с цитоскелетом, а в случае ядерной мембраны с хромосомами. Структурные мембранные белки, как правило, лишены ферментативных свойств (возможно они просто пока мало изучены в химическом отношении). Их исследование затрудняется главным образом двумя обстоятельствами. Во-первых, структурные белки «немы» - не обладают известной ферментативной активностью. Во-вторых, структурные белки имеют в составе своих молекул обширные гидрофобные участки. При очистке они образуют тесные ассоциаты друг с другом или с липидами, что усложняет их изучение.
Нейроспецифический белок В-50 - один из основных фосфорилируемых структурных белков плазматических мембран синаптических контактов. Методами иммунохимии установлено, что он локализован преимущественно в пресинаптических мембранах. Молекулярная масса белка 48 кПа. Он является эндогенным субстратом - зависимой протеинкиназы С. Активаторы протеинкиназы С стимулируют процесс синаптической передачи в срезах гиппокампа. Фосфорилирование белка В-50 приводит к увеличению времени возбужденного состояния синапса, что способствует удержанию ионных каналов в активированном (открытом) состоянии (в некоторых публикациях этот феномен называют состоянием проторенности синапса). Влияние фосфорилированного белка В-50 на метаболизм фосфоинозитидов может быть одной из причин этого феномена. Интересно, что в процессе старения организма интенсивность фосфорилирования белка В-50 в мозге снижается, что, возможно, и обусловливает снижение пластичности синапсов.
Еще одно доказательство роли процессов фосфорилирования белка В-50 в функционировании синапсов получено в экспериментах in vitro, подтвердивших, что нейропептид - фрагмент АКТГ1 _24 - в 10 раз более эффективно тормозит фосфорилирование В-50 в синаптических мембранах из септальной области мозга, чем в мембранах целого мозга.
В группу мембранных белков также входят множество белков-ферментов, образующих ионные каналы, - Na/К- и Са-АТФазы, рецепторные белки, синапсины и др.
Плазмин - сериновая протеиназа, в плазме крови действует в основном как тромболитический фермент, а также деградирует многие компоненты внеклеточного матрикса. В мозге плазмин вовлекается в осуществление многочисленных функций, таких как нейрональная пластичность, обучение и память. Активация плазминогеновой системы наблюдается в мозге во время и в первые дни после инсульта. При болезни Альцгеймера, напротив, происходит снижение уровня плазмина в тканях мозга.
Входящий в состав плазматических мембран нейронов плазмин находится в ассоциации с богатыми холестерином рафтами, которые считаются местом преимущественного образования амилоидного вета-А. Это свидетельствует о наличии функциональной связи между плазмином, холестерином и метаболизмом мозга.
Эндотепинконвертирующий фермент (ЕСЕ-1) является еще одним амилоиддеградирующим ферментом, который на 37% гомологичен НЕП по аминокислотной последовательности.ЕСЕ-1 тоже является мембраносвязанной цинкзависимой металлопротеазой, также как и НЕП он способен расщеплять большое число биологически активных веществ, включая брадикинин, нейротензин, ангиотензин-1 и В-цепь инсулина. В отличие от НЕП, ЕСЕ-1 существует в виде димеров, субъединицы которых соединены дисульфидной связью.
Эти две металлопротеазы (ЕСЕ-1 и НЕП) различаются и по чувствительности к ингибиторам. Для ингибирования НЕП требуются наномолярные концентрации тиорфана и фосфорамидона, в то время как ЕСЕ-1 ингибируется микромолярными концентрациями фосфорамидона и не чувствителен к тиорфану.
ЕСЕ-1 обнаружен во многих органах и тканях. Наиболее обогащены этим ферментом эндотелиальные клетки, он также экспрессируется в нервной ткани и мышцах. В гладкомышечных клетках ЕСЕ-1 находится в комплексе с альфа-актиновыми филаментами.
В настоящее время известны четыре изофоры ЕСЕ-1 человека (1a, 1b, 1c и 1d), которые не имеют существенных каталитических отличий, но различаются по внутриклеточной локализации. Изоформы 1a, 1b, 1c и 1d находятся на поверхности клетки, а ЕСЕ-1 является внутриклеточной формой, локализованной в аппарате Гольджи.
Обнаружен еще один белок, подобный ЕСЕ-1, который локализован преимущественно в мозге и в незначительных количествах - в эндотелиальных и гладкомышечных клетках. Его первичная структура на 59°/о идентична аминокислотной последовательности ЕСЕ-1. Он обозначается как ЕСЕ-2 и отличается от ЕСЕ-1 более кислым рН-оптимумом.
| | | | | | | | | 10 |
Как правило именно белки ответственны за функциональную активность мембран. К ним относятся разнообразные ферменты транспортные белки рецепторы каналы поры и. До этого считалось что мембранные белки имеют исключительно β складчатую структуру вторичная структура белка но данные работы показали что мембраны содержат большое количество α спиралей. Дальнейшие исследования показали что мембранные белки могут глубоко проникать в липидный бислой или даже пронизывать его и их стабилизация осуществляется за счёт гидрофобных...
Поделитесь работой в социальных сетях
Если эта работа Вам не подошла внизу страницы есть список похожих работ. Так же Вы можете воспользоваться кнопкой поиск
Лекция 5
Строение и функции мембранных белков
Клеточные мембраны содержат белка от 20 до 80% (по весу). Как правило, именно белки ответственны за функциональную активность мембран. К ним относятся разнообразные ферменты, транспортные белки, рецепторы, каналы, поры и. т.д., которые обеспечивают уникальность функций каждой мембраны. Первые успехи в изучении мембранных белков были достигнуты тогда, когда биохимики научились использовать детергенты для выделения мембранных белков в функционально активной форме. Это были работы по изучению ферментных комплексов внутренней мембраны митохондрий. До этого считалось, что мембранные белки имеют исключительно β складчатую структуру (вторичная структура белка), но данные работы показали, что мембраны содержат большое количество α спиралей. Значительно реже встречается β спираль, которой, однако, придают важное биологическое значение. Дело в том, что на участках, окружённых липидами, β спираль представляет собой полый цилиндр, в наружной стенке которого сосредоточены неполярные (гидрофобные) аминокислотные остатки, а во внутренней гидрофильные. Такой цилиндр мог бы образовать в мембране канал, через который свободно проходят ионы и водорастворимые вещества. Дальнейшие исследования показали, что мембранные белки могут глубоко проникать в липидный бислой или даже пронизывать его и их стабилизация осуществляется за счёт гидрофобных взаимодействий. Существует, как минимум, четыре вида расположения белков в мембранах: Первый вид трансмембранный, когда белок пронизывает всю мембрану, а гидрофобный участок белка имеет α конфигурацию. Похожее расположение в мембране имеет молекула бактериородопсина из Halobacterium halobium его α спирали последовательно пересекают бислой; Второй вид связывание с помощью гидрофобного якоря, когда у белка есть короткий участок, состоящего из гидрофобных остатков аминокислот вблизи карбоксильного конца. Это, так называемый, гидрофобный якорь, который можно удалить с помощью протеолиза, а высвобождённый белок становится водорастворимым. Такое расположение в мембране присуще многим цитохромам. Третий вид связывание с поверхностью бислоя, когда взаимодействие белков имеет в первую очередь электростатическую природу или гидрофобную природу. Данный тип взаимодействия может использоваться как дополнение к другим взаимодействиям, например, к трансмембранному заякориванию. Четвёртый тип- связывание с белками, погружёнными в бислой, это когда некоторые белки связываются с белками, которые располагаются внутри липидного бислоя. Например, F 1 - часть Н + - АТФазы, которая связывается с F 0 частью, погружённой в мембрану, а также некоторые белки цитоскелета.
В основе современных представлений о структуре мембранных белков лежит идея о том, что их полипептидная цепь уложена так, чтобы образовалась неполярная, гидрофобная поверхность, контактирующая с неполярной областью липидного бислоя. Полярные домены белковой молекулы могут взаимодействовать с полярными головками липидов на поверхности бислоя. Многие белки являются трансмембранными и пронизывают бислой. Некоторые белки, по видимому, связаны с мембраной лишь за счёт их взаимодействия с другими белками.
Многие мембранные белки обычно связываются с мембраной с помощью нековалентных взаимодействий. Однако есть белки, которые связаны с липидами ковалентно. Многие белки плазматических мембран относятся к классу гликопротеинов. Углеводные остатки этих белков всегда находятся с наружной стороны плазматической мембраны.
Обычно мембранные белки подразделяют на наружные (периферические) и внутренние (интегральные). При этом критерием служит степень жёсткости обработки, необходимой для извлечения этих белков из мембраны. Периферические белки высвобождаются при промывании мембран буферными растворами с низкой ионной силой, низким или, наоборот, с высоким значением рН и в присутствии хелатирующих агентов (например, ЭДТА), связывающих двухвалентные катионы. Часто бывает, что очень трудно отличить периферические мембранные белки от белков, связавшихся с мембраной в процессе выделения.
Для высвобождения интегральных мембранных белков необходимо использовать детергенты или даже органические растворители.
Многие мембранные белки эукариот и прокариот ковалентно связаны с липидами, которые присоединяются к полипептиду после трансляции.
Мембранные белки, ковалентно связанные с липидами
(А) Белки, к которым присоединена миристиновая кислота
(Б) Белки, к которым присоединена пальмитиновая кислота
(В) Белки с гликозилфосфатидилинозитольным якорем
4. Адгезивная молекула нервных клеток |
В некоторых случаях эти липиды играют роль гидрофобного якоря, с помощью которого белок прикрепляется к мембране. В других случаях липиды, вероятно, выполняют функцию помощника при миграции белка в соответствующую область клетки или (как в случае белков оболочки вирусов) в слиянии мембран.
У прокариот наиболее полно охарактеризован белок липопротеин Брауна основной липопротеин наружной мембраны E . coli . Зрелая форма этого белка содержит ацилглицерол, который связан тиоэфирной связью с N концевым цистеином. Кроме того, N концевая аминокислота связана с жирной кислотой амидной связью. Мембраносвязанная форма пенициллазы прикрепляется к цитоплазматической мембране с помощью N концевого ацилглицерола аналогично липопротеинам мембраны.
Мембранные белки эукариот ковалентно связанные с липидами, как показано в таблице, их можно разделить на три класса. Белки первых двух классов, по видимому, локализованы в основном на цитоплазматической поверхности плазматической мембраны, а белки третьего класса на наружной поверхности.
Существуют мембранные белки, которые ковалентно связаны с углеводами. К ним относятся поверхностные белки клеток в основном, выполняющих функции транспорта и рецепции. До сих пор неясно, в чём тут дело. Возможно, это связано с тем, что белки нужно сортировать при направлении их к плазматической мембране. Сахарные остатки могут защищать белок от протеолиза или участвовать в узнавании или адгезии. Поэтому сахарные остатки в мембранных гликопротеинах локализованы исключительно на наружной стороне мембраны.
Можно выделить два основных класса олигосахаридных структур мембранных гликопротеинов: 1) N гликозидные олигосахариды, связанные с белками через амидную группу аспаргина; 2) О-гликозидные олигосахариды, связанные через гидроксильные группы серина и треонина. Данный класс олигосахаридов состоит из трёх подклассов.
- Простой или обогащённый маннозой комплекс, в котором олигосахарид содержит маннозу и N ацетилглюкозамин.
- Нормальный комплекс, в котором обогащённый маннозой кор имеет дополнительные боковые ветви, содержащие другие сахаридные остатки, например сиаловую кислоту.
- Большой комплекс, который связан с анионным переносчиком мембраны эритроцитов
Большинство олигосахаридов мембранных гликопротеинов принадлежат к подклассу 1 или2.
Мембранные белки бактерий
Как уже отмечалось выше, белки в цитоплазматической мембране составляют около 50% её поверхности. Примерно 10% мембраны образовано прочно связанными белковолипидными комплексами. Молекула любого встроенного в мембрану белка окружена 45 130 и более липидными молекулами. Около половины свободных липидов связано с периферическими белками мембраны.
Белковый состав цитоплазматической мембраны бактерий более разнообразен, чем липидный. Так, в цитоплазматической мембране E . coli K 12 обнаружено около 120 различных белков. В зависимости от ориентации в мембране и характера связи с липидным бислоем, как уже отмечалось выше, белки делят на интегральные и периферические. К периферическим белкам бактерий можно отнести ряд ферментов таких как, НАДН дегидрогеназа, малатдегидрогеназа и др., а также некоторые белки, которые входят в состав АТФазного комплекса. Этот комплекс представляет собой группу определённым образом расположенных белковых субъединиц, контактирующих с цитоплазмой, периплазматическим пространством и образующих в мембране канал, через который осуществляется переход протона. Участок комплекса, обозначаемый F 1 , погружён в цитоплазму, а и с компоненты участка F 0 гидрофобными сторонами молекул погружены в мембрану. Субъединица b частично погружена в мембрану своей гидрофобной частью и осуществляет связь мембранной и цитоплазматической частей ферментного комплекса, а также связь синтеза АТФ в участке F 1 с протонным потенциалом в мембране. Субъединицы а, b и с обеспечивают протонный канал. Другие компоненты комплекса обеспечивают его структурную и функциональную целостность.
К интегральным белкам E . coli , которые для проявления энзиматической активности необходимы липиды, можно отнести сукцинатдегидрогеназу, цитохром b . Очень интересными свойствами обладает антибиотики грамицидин А, аламетицин, амфотерицин и нистацин. Они при взаимодействии с мембраной бактерий становятся интегральными белками (антибиотики являются полипептидами и макроциклами).
Грамицидин А это гидрофобный пептид, состоящий из 15 L - D -аминокислот. При встраивании в мембрану он образует каналы, которые пропускают одновалентные катионы. Этот канал, который образует грамицидин А охарактеризован наиболее полно. Канал образован двумя молекулами грамицидина А. В результате чередования L - и D - аминокислот образуется спираль, в которой боковые цепи располагаются снаружи, а карбоксильные группы остова внутри канала. Этот тип спирали, не встречается больше ни в каких белках и образуется из за необычного чередования стереоизомеров аминокислот в грамицидине А. Грамицидиновый канал, как уже отмечалось выше, катионселективен. Небольшие неорганические и органические катионы проходят через него, в тоже время проницаемость по Cl - равна нулю.
Аламетицин это пептидный антибиотик из 20 аминокислотных остатков, способный образовывать в мембране электовозбудимые каналы. Аминокислотная последовательность аламетицина включает необычные остатки α аминомасляную кислоту и L фенилаланин. При связывании с мембраной в отличие от грамицидина А он образует пору. Она намного по размеру меньше, чем канал, который образует грамицидин А. Прежде всего это связано с тем, что пространство вокруг α спирали слишком мало, чтобы через него мог пройти ион.
Марколидные антибиотки, такие как, нистатин и амфотерицин связываются с холестерином и образуют каналы. Каналы образуют 8 10 молекул этих полиеновых антибиотиков, через которые, правда, с невысокими скоростями проникают ионы.
Другие похожие работы, которые могут вас заинтересовать.вшм> |
|||
| 21572. | СТРОЕНИЕ И ФУНКЦИИ БЕЛКОВ | 227.74 KB | |
| Содержание белков в организме человека выше чем содержание липидов углеводов. Преобладание в тканях белков по сравнению с другими веществами выявляется при расчёте содержания белков на сухую массу тканей. Содержание белков в различных тканях колеблется в определённом интервале. | |||
| 17723. | Мозжечек, строение и функции | 22.22 KB | |
| 3 Общее строение головного мозга. В нервной системе выделяют также центральную часть ЦНС которая представлена головным и спинным мозгом и периферическую часть в которую входят нервы нервные клетки нервные узлы ганглии и сплетения топографически лежащие вне спинного и головного мозга. Объектом исследования является анатомия головного мозга. Данная цель предмет и объект подразумевают постановку и решение следующих задач: описать общий план строения головного мозга изучить анатомическое строение мозжечка выделить... | |||
| 5955. | Органы растений: их функции, строение и метаморфозы. | 16.94 KB | |
| Органы цветка являются видоизмененными листьями: покровные листья формируют чашелистики и лепестки а спорообразующие листья дают начало тычинкам и пестикам. Побег включает: а стебель б листья в вегетативные почки г цветки д плоды. Стеблем называется вегетативный орган растения который выполняет многочисленные функции: несёт листья или тяжелую крону из ветвей и листьев; связывает корни и листья; на нем образуются цветки; по нему передвигается вода с минеральными веществами и органическими соединениями; молодые стебли... | |||
| 5067. | Гладкие мышцы. Строение, функции, механизм сокращения | 134.79 KB | |
| Мышцы или мускулы от лат. Мышцы позволяют двигать частями тела и выражать в действиях мысли и чувства. Гладкие мышцы являются составной частью некоторых внутренних органов и участвуют в обеспечении функции выполняемые этими органами. | |||
| 6233. | Строение и функции ядра. Морфология и химический состав ядра | 10.22 KB | |
| От цитоплазмы ядра обычно отделяются четкой границей. Бактерии и синезеленые водоросли не имеют сформированного ядра: их ядро лишено ядрышка не отделено от цитоплазмы отчетливо выраженной ядерной мембраной и носит название нуклеоид. Форма ядра. | |||
| 9495. | Классификация, характеристика ассортимента пушно-мехового сырья и пушно-мехового полуфабриката, строение пушно-меховой шкуры, строение волоса и разновидность его форм, технология изготовления пушнины | 1.05 MB | |
| Меховые пластины полосы определенной формы сшитые из подобранных выделанных шкурок и предназначенные для раскроя на детали меховых изделий. К зимним видам пушного сырья относятся шкурки и шкуры пушных зверей добыча которых производится преимущественно в зимнее время когда качество шкурок особенно высоко. СТРОЕНИЕ И ХИМИЧЕСКИЙ СОСТАВ ШКУРОК ПУШНОМЕХОВОГО и овчинношубного СЫРЬЯ ПОНЯТИЕ О ТОПОГРАФИИ ШКУРКИ Шкуркой называют наружный покров животного отделенный от его тушки и состоящий из кожной ткани и волосяного покрова. У... | |||
| 8011. | Свойства мембранных липидов | 10.13 KB | |
| Некоторые липиды способствуют стабилизации сильно искривлённых участков мембраны образованию контакта между мембранами или связыванию определённых белков поскольку форма этих молекул благоприятствует нужной упаковке бислоя на соответствующих участках мембраны. Под жидкостным состоянием понимают способность фосфолипидных молекул к вращению и латеральному перемещению в соответствующем лепестке мембраны. Они вытянуты и ориентированы перпендикулярно плоскости мембраны. В состоянии жидкого кристалла молекулы жирных кислот подвижны но... | |||
| 8014. | Химический состав мембранных липидов | 10.81 KB | |
| Прежде всего это связано с множеством функций которые выполняют липиды в мембранах. Фосфатидная кислота в свободном виде содержится в мембранах бактерий в небольшом количестве обычно же к ней присоединены остатки спиртов аминокислот и др. Эти липиды являются сложными эфирами жирных кислот и глицерола и широко представлены во многих мембранах эукариотических и прокариотических клеток за исключением архебактерий. Они содержатся в большом количестве во внутренней мембране митохондрий в мембране хлоропластов и в некоторых бактериальных... | |||
| 21479. | ОБМЕН БЕЛКОВ | 150.03 KB | |
| Различают три вида азотистого баланса: азотистое равновесие положительный азотистый баланс отрицательный азотистый баланс При положительном азотистом балансе поступление азота преобладает над его выделением. При заболевании почек возможен ложный положительный азотистый баланс при котором происходит задержка в организме конечных продуктов азотистого обмена. При отрицательном азотистом балансе преобладает выделение азота над его поступлением. Это состояние возможно при таких заболеваниях как туберкулез ревматизм онкологические... | |||
| 15073. | Рассмотрение мембранных (ионоселективных) электродов с различного рода мембранами | 127.48 KB | |
| Для этого существуют разнообразные ионоселективные электроды главной особенностью которых является так называемая селективность к определенному виду ионов. Электроды с жидкой и пленочной мембраной Жидкие мембраны это растворы в органических растворителях ионообменных веществ жидкие катиониты или аниониты или нейтральных хелатов отделенные от водных растворов нейтральными пористыми перегородками полимерными стеклянными или др. В настоящее время промышленность выпускает пленочные ионоселективные электроды на катионы N К NH4 Са2... | |||
Биологическая химия Лелевич Владимир Валерьянович
Белки мембран.
Белки мембран.
Мембранные белки отвечают за функциональную активность мембран и на их долю приходится от 30 до 70%. Белки мембран отличаются по своему положению в мембране. Они могут глубоко проникать в липидный бислой или даже пронизывать его – интегральные белки, разными способами прикрепляться к мембране – поверхностные белки, либо, ковалентно контактировать с ней – заякоренные белки. Поверхностные белки почти всегда гликозилированы. Олигосахаридные остатки защищают белок от протеолиза, участвуют в узнавании лигандов и адгезии.
Белки, локализованные в мембране, выполняют структурную и специфические функции:
1. транспортную;
2. ферментативную;
3. рецепторную;
4. антигенную.
Из книги Заводи кого угодно, только НЕ КРОКОДИЛА! автора Орсаг МихайНу а белки? В шестидесятых годах я неоднократно пытался завести в доме и белок, но каждая такая попытка кончалась самым печальным образом. Через некоторое время белки слабели, задние конечности у них отнимались и несчастные животные в судорогах погибали. Поначалу я
Из книги Геном человека: Энциклопедия, написанная четырьмя буквами автора Из книги Геном человека [Энциклопедия, написанная четырьмя буквами] автора Тарантул Вячеслав ЗалмановичНе все гены кодируют белки Прежде всего, следует отметить, что кроме генов, кодирующих белки, в геноме имеются еще гены, на которых синтезируется РНК, которые не являются мРНК (то есть, не кодируют белок), но выполняют ряд самостоятельных важных функций в клетках. В
Из книги Живые часы автора Уорд Ритчи11. Белки в колесе Чтобы познакомиться с современными поисками решения проблемы живых часов, обратимся к исследованиям, проводившимся биологами в последние годы. В числе первых следует, пожалуй, назвать работу Патриции де Курси.В 1955 году де Курси получила диплом
Из книги Биология [Полный справочник для подготовки к ЕГЭ] автора Лернер Георгий Исаакович Из книги Кровь: река жизни [От древних легенд до научных открытий] автора Азимов АйзекГлава 11 Эти подвижные белки В начале предыдущей главы я упомянул, что органические компоненты пищи делятся на три группы. Я рассказал об одной из этих групп: углеводах. Далее логично было бы перейти к белкам, потому что их метаболизм в организме происходит параллельно с
Из книги Логика случая [О природе и происхождении биологической эволюции] автора Кунин Евгений ВикторовичГлава 12 Происхождение жизни. Возникновение трансляции, репликации, метаболизма и мембран: биологический, геохимический и космологический подходы Пер. А. НеизвестногоВ предыдущей главе мы обсудили возможные сценарии возникновения клеток и (будем надеяться) достигли
Из книги Гены и развитие организма автора Нейфах Александр Александрович2. Белки хроматина Мы уже знаем, что хроматин состоит из ДНК и гистонов в равном весовом количестве и негистоновых белков (НГБ), которых в неактивных районах хромосомы всего 0,2 веса ДНК, а в активных - более чем 1,2 (в среднем НГБ мепьше, чем ДНК). Мы знаем также, что гистоны
Из книги Мир животных автора Ситников Виталий Павлович Из книги Современное состояние биосферы и экологическая политика автора Колесник Ю. А.4.1. Образование мембран – основа начала жизни Рассматривая гигантское многообразие современных живых организмов, можно прийти к выводу, что существовало множество путей развития, берущих начало от реликтовых форм жизни. На самом деле исследования молекулярной эволюции
Из книги Секреты наследственности человека автора Афонькин Сергей ЮрьевичКлетки, белки и гены Жизнь есть способ существования белковых тел. Ф. Энгельс Наше тело является империей клеток, каждая из которых представляет собой миниатюрную фабрику для производства белков. Многие из этих важнейших макромолекул могут быть выделены из организма в
Из книги Антропология и концепции биологии автора Курчанов Николай АнатольевичБелки Белки имеют первостепенное значение в жизни организмов. Огромное разнообразие живых существ в значительной степени определяется различиями в составе имеющихся в их организме белков. Например, в организме человека их известно более 5 млн.Белки – это полимеры,
Из книги Биологическая химия автора Лелевич Владимир ВалерьяновичХимический состав мембран. Мембраны состоят из липидных и белковых молекул, относительное количество которых у разных мембран широко колеблется. Углеводы содержатся в форме гликопротеинов, гликолипидов и составляют 0,5%-10% веществ мембраны. Согласно жидкостно-мозаичной
Из книги автораЛипиды мембран. Мембранные липиды – амфифильные молекулы, т.е. в молекуле есть как гидрофильные группы (полярные головки), так и алифатические радикалы (гидрофобные хвосты), самопроизвольно формирующие бислой, в котором хвосты липидов обращены друг к другу. Толщина
Из книги автораБелки Пищевая ценность белка обеспечивается наличием незаменимых аминокислот, углеводородные скелеты которых не могут синтезироваться в организме человека, и они соответственно должны поступать с пищей. Они также являются основными источниками азота. Суточная
Из книги автораБелки мышечной ткани Выделяют три группы белков:1. миофибриллярные белки – 45 %;2. саркоплазматические белки – 35 %;3. белки стромы – 20 %.Миофибриллярные белки.К этой группе относятся:1. миозин;2. актин;3. актомиозин;а также так называемые регуляторные белки:4. тропомиозин;5.