Vee hulk erinevates rakkudes. Taimeraku keemiline koostis. Uue materjali õppimine
Veesisaldus erinevates taimeorganites varieerub üsna suurtes piirides. See varieerub sõltuvalt keskkonnatingimustest, taimede vanusest ja tüübist. Seega on salatilehtede veesisaldus 93-95%, mais - 75-77%. Vee hulk ei ole taimede erinevates organites ühesugune: päevalille lehed sisaldavad 80-83% vett, varred - 87-89%, juured - 73-75%. Veesisaldus, mis on võrdne 6-11%, on tüüpiline peamiselt õhu käes kuivanud seemnetele, milles elutähtsad protsessid on pärsitud.
Vesi sisaldub elusrakkudes, ksüleemi surnud elementides ja rakkudevahelistes ruumides. Rakkudevahelistes ruumides on vesi aurulises olekus. Lehed on taime peamised aurustusorganid. Sellega seoses on loomulik, et suurim kogus vett täidab lehtede rakkudevahelised ruumid. Vedelas olekus leidub vett raku erinevates osades: rakumembraanis, vakuoolides ja protoplasmas. Vakuoolid on raku kõige veerikkam osa, kus selle sisaldus ulatub 98%-ni. Suurima veesisalduse korral on veesisaldus protoplasmas 95%. Madalaim veesisaldus on iseloomulik rakumembraanidele. Veesisalduse kvantitatiivne määramine rakumembraanides on keeruline; ilmselt jääb see vahemikku 30–50%.
Erinevad on ka vee vormid taimeraku erinevates osades. Vaakumrakkude mahlas domineerib suhteliselt madala molekulmassiga ühendite (osmootselt seotud) peetav vesi ja vaba vesi. Taimeraku kestas seovad vett peamiselt kõrgpolümeersed ühendid (tselluloos, hemitselluloos, pektiinained), st kolloidselt seotud vesi. Tsütoplasmas endas on kolloidselt ja osmootselt seotud vaba vesi. Valgumolekuli pinnast kuni 1 nm kaugusel asuv vesi on kindlalt seotud ja sellel puudub korrapärane kuusnurkne struktuur (kolloidseotud vesi). Lisaks on protoplasmas teatud kogus ioone ja järelikult on osa veest osmootselt seotud.
Vaba ja seotud vee füsioloogiline tähtsus on erinev. Enamik teadlasi arvab, et füsioloogiliste protsesside intensiivsus, sealhulgas kasvukiirused, sõltub eelkõige vaba vee sisaldusest. Seotud vee sisalduse ja taimede vastupidavuse vahel ebasoodsatele välistingimustele on otsene seos. Neid füsioloogilisi seoseid ei täheldata alati.
Taimerakk imab vett vastavalt osmoosiseadustele. Osmoosi täheldatakse kahe erineva ainete kontsentratsiooniga süsteemi juuresolekul, kui need suhtlevad poolläbilaskva membraaniga. Sel juhul võrdsustuvad kontsentratsioonid vastavalt termodünaamika seadustele aine tõttu, mille jaoks membraan on läbilaskev.
Kui vaadelda kahte erineva osmootselt aktiivsete ainete kontsentratsiooniga süsteemi, siis järeldub, et kontsentratsioonide võrdsustamine süsteemides 1 ja 2 on võimalik ainult tänu vee liikumisele. Süsteemis 1 on vee kontsentratsioon suurem, seega suunatakse veevool süsteemist 1 süsteemi 2. Kui tasakaal on saavutatud, on tegelik vooluhulk null.
Taimerakku võib pidada osmootseks süsteemiks. Rakku ümbritsev rakusein on teatud elastsusega ja venitatav. Vakuooli kogunevad osmootse aktiivsusega vees lahustuvad ained (suhkrud, orgaanilised happed, soolad). Tonoplast ja plasmalemma täidavad selles süsteemis poolläbilaskva membraani funktsiooni, kuna need struktuurid on selektiivselt läbilaskvad ja vesi läbib neid palju kergemini kui rakumahlas ja tsütoplasmas lahustunud ained. Sellega seoses, kui rakk satub keskkonda, kus osmootselt aktiivsete ainete kontsentratsioon on väiksem kui kontsentratsioon rakus (või rakk asetatakse vette), peab vesi osmoosiseaduste kohaselt rakku sisenema. .
Veemolekulide võimet liikuda ühest kohast teise mõõdetakse veepotentsiaaliga (Ψw). Termodünaamika seaduste kohaselt liigub vesi alati kõrgema veepotentsiaaliga alalt madalama potentsiaaliga piirkonda.
Vee potentsiaal(Ψ в) on vee termodünaamilise oleku indikaator. Veemolekulidel on kineetiline energia, nad liiguvad vedelikus ja veeaurus juhuslikult. Veepotentsiaal on suurem süsteemis, kus molekulide kontsentratsioon on suurem ja nende kogukineetiline energia suurem. Puhas (destilleeritud) vesi omab maksimaalset veepotentsiaali. Sellise süsteemi veepotentsiaal võetakse tinglikult nulliks.
Veepotentsiaali ühikud on rõhu ühikud: atmosfäär, paskalid, baarid:
1 Pa = 1 N/m 2 (N-newton); 1 bar = 0,987 atm = 10 5 Pa=100 kPa;
1 atm = 1,0132 baari; 1000 kPa = 1 MPa
Teise aine lahustamisel vees väheneb vee kontsentratsioon, veemolekulide kineetiline energia ja veepotentsiaal väheneb. Kõigis lahendustes on vee potentsiaal väiksem kui puhtal veel, s.t. standardtingimustes väljendatakse seda negatiivse väärtusena. Kvantitatiivselt väljendatakse seda vähenemist kogusega, mida nimetatakse osmootne potentsiaal(Ψ osm.). Osmootne potentsiaal on lahustunud ainete olemasolu tõttu veepotentsiaali vähenemise mõõt. Mida rohkem on lahuses lahustunud molekule, seda väiksem on osmootne potentsiaal.
Kui vesi siseneb rakku, suureneb selle suurus, suureneb rakusisene hüdrostaatiline rõhk, mis sunnib plasmalemma suruma vastu rakuseina. Rakusein avaldab omakorda vasturõhku, mida iseloomustab survepotentsiaal(Ψ rõhk) või hüdrostaatiline potentsiaal, on see tavaliselt positiivne ja mida suurem, seda rohkem vett rakus.
Seega sõltub raku veepotentsiaal osmootselt aktiivsete ainete kontsentratsioonist - osmootsest potentsiaalist (Ψ osm.) Ja rõhupotentsiaalist (Ψ rõhk).
Eeldusel, et vesi ei suru rakumembraanile (plasmolüüsi või närbumise seisund), on rakumembraani vasturõhk null, vee potentsiaal on võrdne osmootse:
Ψ sisse. = Ψ osm.
Kui vesi rakku siseneb, ilmneb rakumembraani vasturõhk, veepotentsiaal võrdub osmootse potentsiaali ja rõhupotentsiaali vahega:
Ψ sisse. = Ψ osm. + Ψ rõhk
Erinevus rakumahla osmootse potentsiaali ja rakumembraani vasturõhu vahel määrab vee voolu igal hetkel.
Tingimusel, et rakumembraan on piirini venitatud, tasakaalustab osmootne potentsiaal täielikult rakumembraani vasturõhuga, veepotentsiaal muutub nulliks ja vesi lakkab rakku voolamast:
- Ψ osm. = Ψ rõhk , Ψ c. = 0
Vesi voolab alati negatiivsema veepotentsiaali suunas: süsteemist, kus energia on suurem, süsteemi, kus energiat on vähem.
Vesi võib rakku sattuda ka paisumisjõudude toimel. Valgud ja muud rakku moodustavad ained, millel on positiivselt ja negatiivselt laetud rühmad, tõmbavad ligi veedipoole. Hemitselluloose ja pektiinaineid sisaldav rakusein ning tsütoplasma, milles kõrgmolekulaarsed polaarsed ühendid moodustavad umbes 80% kuivmassist, on võimelised paisuma. Vesi tungib paisumisstruktuuri difusiooni teel, vee liikumine järgib kontsentratsioonigradienti. Turse jõudu tähistatakse terminiga maatriksi potentsiaal(Ψ mat.). See sõltub raku suure molekulmassiga komponentide olemasolust. Maatriksi potentsiaal on alati negatiivne. Suur tähtsusΨ mat. on siis, kui vett neelavad struktuurid, milles puuduvad vakuoolid (seemned, meristeemrakud).
Vesi on kõige levinum ühend Maal ja elusorganismides. Veesisaldus rakkudes oleneb ainevahetusprotsesside iseloomust: mida intensiivsemad need on, seda suurem on veesisaldus.
Täiskasvanud inimese rakud sisaldavad keskmiselt 60–70% vett. 20% vee kadumisel organismid surevad. Ilma veeta võib inimene elada kuni 7 päeva, ilma toiduta aga mitte rohkem kui 40 päeva.

Riis. 4.1. Veemolekuli (H 2 O) ruumiline struktuur ja vesiniksideme teke
Veemolekul (H 2 O) koosneb kahest vesinikuaatomist, mis on kovalentselt seotud hapnikuaatomitega. Molekul on polaarne, kuna see on nurga all painutatud ja hapnikuaatomi tuum tõmbab jagatud elektronid selle nurga alla, nii et hapnik omandab osalise negatiivse laengu ja avatud otstes olevad vesinikuaatomid muutuvad osaliselt positiivseteks laenguteks. Veemolekule saab üksteise külge meelitada positiivsete ja negatiivsete laengutega, moodustades vesinikside (Joon.4.1.).
Tänu ainulaadne struktuur veemolekulid ja nende võime omavahel vesiniksidemeid kasutades siduda, on veel mitmeid omadusi, mis määravad ära selle olulise rolli rakus ja organismis.
Vesiniksidemed põhjustavad suhteliselt kõrgeid keemis- ja aurustumistemperatuure, vee kõrget soojusmahtuvust ja soojusjuhtivust ning universaalse lahusti omadust.
Vesiniksidemed on 15-20 korda nõrgemad kui kovalentsed sidemed. Vedelas olekus vesiniksidemed kas tekivad või katkevad, mis põhjustab veemolekulide liikumise, selle voolavuse.
Vesi määrab raku füüsikalised omadused – mahu, elastsuse (turgor). Rakk sisaldab 95-96% vaba vett ja 4-5% seotust. Seotud vesi moodustab teatud ühendite (näiteks valkude) ümber vesised (solvaatsed) kestad, takistades nende omavahelist koostoimet.
tasuta vesi on hea lahusti paljude anorgaaniliste ja orgaaniliste polaarsete ainete jaoks. Vees hästi lahustuvaid aineid nimetatakse hüdrofiilsed. Näiteks alkoholid, happed, gaasid, enamik naatriumi-, kaaliumisooli jne. Hüdrofiilsete ainete puhul on nende aatomite vaheline sidumisenergia väiksem kui nende aatomite veemolekulide külgetõmbeenergia. Seetõttu on nende molekulid või ioonid kergesti integreeritavad vee üldisesse vesiniksidemete süsteemi.
Vesi kui universaalne lahusti mängib äärmiselt olulist rolli, kuna enamik keemilised reaktsioonid esineb vesilahustes. Ainete tungimine rakku ja jääkainete eemaldamine sellest on enamikul juhtudel võimalik ainult lahustunud kujul.
Vesi ei lahusta mittepolaarseid (laenguta) aineid, kuna ei saa nendega moodustada vesiniksidemeid. Vees lahustumatud aineid nimetatakse hüdrofoobne . Nende hulka kuuluvad rasvad, rasvataolised ained, polüsahhariidid, kumm.
Mõnel orgaanilisel molekulil on kaks omadust: mõnes piirkonnas on need polaarsed rühmad ja teistes - mittepolaarsed. Selliseid aineid nimetatakse amfipaatiline või amfifiilne. Nende hulka kuuluvad valgud, rasvhapped, fosfolipiidid, nukleiinhapped. Amfifiilsed ühendid mängivad olulist rolli bioloogiliste membraanide, keerukate supramolekulaarsete struktuuride organiseerimisel.
Vesi on reaktsioonides otseselt seotud hüdrolüüs- orgaaniliste ühendite lagunemine. Samal ajal lisatakse spetsiaalsete ensüümide toimel orgaaniliste molekulide vabadele valentsidele OH ioone. - ja H + vesi. Selle tulemusena moodustavad nad uusi aineid, millel on uued omadused.
Vesi on suure soojusmahutavusega (s.t. võime absorbeerida soojust oma temperatuuri väikeste muutustega) ja hea soojusjuhtivusega. Tänu nendele omadustele hoitakse rakus (ja kehas) temperatuur teatud tasemel koos ümbritseva keskkonna temperatuuri oluliste muutustega.
Tähtis bioloogiline tähtsus taimede funktsioneerimiseks on külmaverelistel loomadel asjaolu, et lahustunud ainete (süsivesikud, glütserool) mõjul võib vesi muuta oma omadusi, eelkõige külmumis- ja keemistemperatuure.
Vee omadused on elusorganismide jaoks nii olulised, et meie teadaoleva elu olemasolu pole võimalik ette kujutada mitte ainult Maal, vaid ka igal teisel planeedil ilma piisava veevarudeta.
MINERAALSOOL
Need võivad olla lahustunud või lahustumata olekus. Mineraalsoolade molekulid vesilahuses lagunevad katioonideks ja anioonideks.
Veesisaldus erinevates taimeorganites varieerub üsna suurtes piirides. See varieerub sõltuvalt keskkonnatingimustest, taimede vanusest ja tüübist. Seega on salatilehtede veesisaldus 93-95%, mais - 75-77%. Vee hulk ei ole taimede erinevates organites ühesugune: päevalille lehed sisaldavad 80-83% vett, varred - 87-89%, juured - 73-75%. Veesisaldus, mis on võrdne 6-11%, on tüüpiline peamiselt õhu käes kuivanud seemnetele, milles elutähtsad protsessid on pärsitud.
Vesi sisaldub elusrakkudes, ksüleemi surnud elementides ja rakkudevahelistes ruumides. Rakkudevahelistes ruumides on vesi aurulises olekus. Lehed on taime peamised aurustusorganid. Sellega seoses on loomulik, et suurim kogus vett täidab lehtede rakkudevahelised ruumid. Vedelas olekus leidub vett raku erinevates osades: rakumembraanis, vakuoolis, tsütoplasmas. Vakuoolid on raku kõige veerikkam osa, kus selle sisaldus ulatub 98%-ni. Suurima veesisalduse korral on veesisaldus tsütoplasmas 95%. Madalaim veesisaldus on iseloomulik rakumembraanidele. Veesisalduse kvantitatiivne määramine rakumembraanides on keeruline; ilmselt jääb see vahemikku 30–50%.
Erinevad on ka vee vormid taimeraku erinevates osades. Vaakumrakkude mahlas domineerib suhteliselt madala molekulmassiga ühendite (osmootselt seotud) peetav vesi ja vaba vesi. Taimeraku kestas seovad vett peamiselt kõrgpolümeersed ühendid (tselluloos, hemitselluloos, pektiinained), st kolloidselt seotud vesi. Tsütoplasmas endas on kolloidselt ja osmootselt seotud vaba vesi. Valgumolekuli pinnast kuni 1 nm kaugusel asuv vesi on kindlalt seotud ja sellel puudub korrapärane kuusnurkne struktuur (kolloidseotud vesi). Lisaks on tsütoplasmas teatud kogus ioone ja järelikult on osa veest osmootselt seotud.
Vaba ja seotud vee füsioloogiline tähtsus on erinev. Enamiku teadlaste arvates sõltub füsioloogiliste protsesside intensiivsus, sealhulgas kasvukiirused, eelkõige vaba vee sisaldusest. Seotud vee sisalduse ja taimede vastupidavuse vahel ebasoodsatele välistingimustele on otsene seos. Neid füsioloogilisi seoseid ei täheldata alati.
Normaalseks eksisteerimiseks peavad rakud ja taimeorganism tervikuna sisaldama teatud kogust vett. See on aga kergesti teostatav ainult vees kasvavate taimede puhul. Maismaataimede jaoks muudab selle ülesande keeruliseks asjaolu, et aurustumisprotsessis kaob taimeorganismis pidevalt vett. Vee aurustumine taime poolt saavutab tohutud mõõtmed. Võib tuua näite: üks maisitaim aurustab kasvuperioodil kuni 180 kg vett, aastal 1 hektar metsa. Lõuna-Ameerika aurustab keskmiselt 75 tuhat kg vett päevas. Tohutu veetarbimine on tingitud asjaolust, et enamikul taimedel on märkimisväärne lehepind, mis asub veeauruga küllastumata atmosfääris. Samas on ekstensiivse lehepinna arendamine vajalik ja pika evolutsiooni käigus välja kujunenud, et tagada normaalne toitumine õhus ebaolulises kontsentratsioonis (0,03%) sisalduva süsihappegaasiga. Oma kuulsas raamatus "Taimede võitlus põuaga" K.A. Timirjazev tõi välja vastuolu jäädvustamise vajaduse vahel süsinikdioksiid ja veetarbimise vähendamine jättis jälje kogu taimeorganismi struktuuri.
Aurustumisel tekkiva veekadu kompenseerimiseks peab suur kogus seda pidevalt seadmesse jõudma. Nimetatakse kahte tehases pidevalt toimuvat protsessi – vee sissevoolu ja aurustumist taimede vee tasakaal. Taimede normaalseks kasvuks ja arenguks on vajalik, et veekulu vastaks ligikaudu sissetulekule ehk teisisõnu, et taim vähendaks oma veebilanssi ilma suure puudujäägita. Selleks töötas taim loodusliku valiku protsessis välja kohandused vee neeldumiseks (kolossaalselt arenenud juurestik), vee liikumiseks (spetsiaalne juhtiv süsteem) ja aurustumise vähendamiseks (integreeritud kudede süsteem ja stomataalsete avade automaatse sulgemise süsteem).
Kõigist nendest kohandustest hoolimata täheldatakse taimes sageli veepuudust, st vee tarbimist ei tasakaalusta selle tarbimine transpiratsiooniprotsessis.
Füsioloogilised häired esinevad erinevates taimedes, mille veepuudus on erinev. On taimi, millel on evolutsiooni käigus välja kujunenud mitmesugused kohandused, et taluda dehüdratsiooni (põuakindlad taimed). Taimede veepuuduse vastupanuvõimet määravate füsioloogiliste tunnuste väljaselgitamine on kõige olulisem ülesanne, mille lahendamisel on suur mitte ainult teoreetiline, vaid ka põllumajanduslik praktiline tähtsus. Samas on selle lahendamiseks vajalikud teadmised taimeorganismi veevahetuse kõikidest aspektidest.
AT maakoor leitud umbes 100 keemilised elemendid, kuid neist on eluks vaja vaid 16. Taimeorganismides on kõige levinumad neli elementi – vesinik, süsinik, hapnik, lämmastik, mis tekivad erinevaid aineid. Taimeraku põhikomponendid on vesi, orgaanilised ja mineraalained.
Vesi- elu alus. Veesisaldus taimerakkudes on vahemikus 90–10%. Ta on ainulaadne aine tänu oma keemilisele ja füüsikalised omadused. Vesi on vajalik fotosünteesi protsessiks, ainete transpordiks, rakkude kasvuks, see on paljude biokeemiliste reaktsioonide keskkond, universaalne lahusti jne.
Mineraalid (tuhk)- ained, mis jäävad pärast elunditüki põletamist. Tuhaelementide sisaldus jääb vahemikku 1% kuni 12% kuivkaalust. Peaaegu kõik elemendid, millest koosneb vesi ja muld, leidub taimes. Levinumad on kaalium, kaltsium, magneesium, raud, räni, väävel, fosfor, lämmastik (makroelemendid) ja vask, alumiinium, kloor, molübdeen, boor, tsink, liitium, kuld (mikroelemendid). Mineraalidel on oluline roll rakkude elus – need on osa aminohapetest, ensüümidest, ATP-st, elektronide transpordiahelatest, on vajalikud membraanide stabiliseerimiseks, osalevad ainevahetusprotsessides jne.
orgaaniline aine taimerakud jagunevad: 1) süsivesikuteks, 2) valkudeks, 3) lipiidideks, 4) nukleiinhapeteks, 5) vitamiinideks, 6) fütohormoonideks, 7) sekundaarse ainevahetuse saadusteks.
Süsivesikud moodustavad kuni 90% taimeraku moodustavatest ainetest. Eristama:
Monosahhariidid (glükoos, fruktoos). Monosahhariidid tekivad lehtedes fotosünteesi käigus ja muutuvad kergesti tärkliseks. Nad kogunevad viljadesse, harvemini vartesse, sibulatesse. Monosahhariidid transporditakse rakust rakku. Nad on energiamaterjal, osalevad glükosiidide moodustumisel.
Disahhariidid (sahharoos, maltoos, laktoos jne) tekivad kahest monosahhariidi osakesest. Nad kogunevad juurtesse ja viljadesse.
Polüsahhariidid on taimerakkudes väga laialt levinud polümeerid. Sellesse ainete rühma kuuluvad tärklis, inuliin, tselluloos, hemitselluloos, pektiin, kalloosi.
Tärklis on taimeraku peamine säilitusaine. Primaarne tärklis moodustub kloroplastides. Taime rohelistes osades jaguneb see mono- ja disahhariidideks ning transporditakse mööda veenide floeemi taime kasvuosadesse ja säilitusorganitesse. Säilitusorganite leukoplastides sünteesitakse sahharoosist tärkliseteradena sekundaarne tärklis.
Tärklise molekul koosneb amüloosist ja amülopektiinist. Lineaarsed amüloosiahelad, mis koosnevad mitmest tuhandest glükoosijäägist, on võimelised spiraalselt hargnema ja seega võtma kompaktsema kuju. Hargnenud polüsahhariidis amülopektiinis tagab kompaktsuse 1,6-glükosiidsidemete moodustumise tõttu intensiivne ahela hargnemine. Amülopektiin sisaldab ligikaudu kaks korda rohkem glükoosijääke kui amüloos.
Lugoli lahusega annab amüloosi vesisuspensioon tumesinise värvi, amülopektiini suspensioon - punakasvioletne, tärklise suspensioon - sinakasvioletne.
Inuliin on fruktoosi polümeer, astrite perekonda kuuluv süsivesik. Seda leidub rakkudes lahustunud kujul. Ei määri joodilahusega, värvib β-naftooliga punaseks.
Tselluloos on glükoosi polümeer. Tselluloos sisaldab umbes 50% taimes leiduvast süsinikust. See polüsahhariid on rakuseina peamine materjal. Tselluloosi molekulid on pikad glükoosijääkide ahelad. Igast ahelast ulatub välja palju OH-rühmi. Need rühmad on suunatud igas suunas ja moodustavad naaberahelatega vesiniksidemeid, mis tagab kõigi ahelate jäiga ristsidumise. Ahelad kombineeritakse omavahel, moodustades mikrofibrillid ja viimased ühendatakse suuremateks struktuurideks - makrofibrillid. Selle konstruktsiooni tõmbetugevus on väga kõrge. Kihtides paiknevad makrofibrillid on sukeldatud tsementeerivasse maatriksisse, mis koosneb pektiinainetest ja hemitselluloosidest.
Tselluloos ei lahustu vees, joodilahusega annab kollase värvuse.
Pektiinid koosnevad galaktoosist ja galakturoonhappest. Pektiinhape on polügalakturoonhape. Need on osa rakuseina maatriksist ja tagavad selle elastsuse. Pektiinid moodustavad keskmise kihi aluse, mis moodustub rakkude vahel pärast jagunemist. Vormi geelid.
Hemitselluloosid on segakoostise makromolekulaarsed ühendid. Need on osa rakuseina maatriksist. Nad ei lahustu vees, hüdrolüüsivad happelises keskkonnas.
Kalloos on glükoosi amorfne polümeer, mida leidub taime keha erinevates osades. Kalloos moodustub floeemi sõelatorudes ja sünteesitakse ka vastusena kahjustustele või kahjulikele mõjudele.
Agar-agar on kõrge molekulmassiga polüsahhariid, mida leidub merevetikates. See lahustub kuumas vees ja pärast jahutamist kõveneb.
Oravad aminohapetest koosnevad makromolekulaarsed ühendid. Elementaarne koostis - C, O, N, S, P.
Taimed on võimelised sünteesima kõiki aminohappeid lihtsamatest ainetest. 20 aluselist aminohapet moodustavad terve hulga valke.
Valkude struktuuri keerukus ja nende funktsioonide äärmuslik mitmekesisus muudavad valkude ühe ja selge klassifikatsiooni loomise ühelgi alusel keeruliseks. Koostise järgi jagunevad valgud lihtsateks ja keerukateks. Lihtne – koosneb ainult aminohapetest, kompleks – koosneb aminohapetest ja mittevalgulisest materjalist (proteesirühm).
Lihtsate valkude hulka kuuluvad albumiinid, globuliinid, histoonid, prolamiinid ja gluteniinid. Albumiinid on vees lahustuvad neutraalsed valgud, mida leidub taimedes harva. Globuliinid on neutraalsed valgud, mis ei lahustu vees, lahustuvad lahjendatud soolalahustes, jaotuvad taimede seemnetes, juurtes, vartes. Histoonid on vees lahustuvad neutraalsed valgud, mis paiknevad kõigi elusrakkude tuumades. Proamiinid - lahustuvad 60-80% etanoolis, leidub teravilja terades. Gluteniinid lahustuvad leeliselahustes, neid leidub teravilja terades, rohelistes taimeosades.
Komplekssete hulka kuuluvad fosfoproteiinid (proteesirühm on fosforhape), lükoproteiinid (süsivesikud), nukleoproteiinid (nukleiinhape), kromoproteiinid (pigment), lipoproteiinid (lipiid), flavoproteiinid (FAD), metalloproteiinid (metall).
Valgud mängivad taimeorganismi elus olulist rolli ning valgud jagunevad olenevalt täidetavast funktsioonist struktuurvalkudeks, ensüümideks, transportvalkudeks, kontraktiilseteks valkudeks, säilitusvalkudeks.
Lipiidid- orgaanilised ained, mis ei lahustu vees ja lahustuvad orgaanilistes lahustites (eeter, kloroform, benseen). Lipiidid jagunevad tõelisteks rasvadeks ja lipoidideks.
Tõelised rasvad on rasvhapete ja teatud tüüpi alkoholi estrid. Need moodustavad vees emulsiooni, leelistega kuumutamisel hüdrolüüsivad. Need on varuained, kogunevad seemnetesse.
Lipoidid on rasvataolised ained. Nende hulka kuuluvad fosfolipiidid (need on osa membraanidest), vahad (moodustavad lehtedele ja viljadele kaitsva katte), steroolid (nad on osa protoplasmast, osalevad sekundaarsete metaboliitide moodustumisel), karotenoidid (punased ja kollased pigmendid, mis on vajalikud kaitsta klorofülli, värvida vilju, õisi), klorofülli (fotosünteesi peamine pigment)
Nukleiinhapped - kõigi elusorganismide geneetiline materjal. Nukleiinhapped (DNA ja RNA) koosnevad monomeeridest, mida nimetatakse nukleotiidideks. Nukleotiidmolekul koosneb viiest süsinikust koosnevast suhkrust, lämmastikalusest ja fosforhappest.
vitamiinid- mitmesuguse keemilise koostisega komplekssed orgaanilised ained. Neil on kõrge füsioloogiline aktiivsus – need on vajalikud valkude, rasvade sünteesiks, ensüümide tööks jne. Vitamiinid jagunevad rasvlahustuvateks ja veeslahustuvateks. Rasvlahustuvate vitamiinide hulka kuuluvad vitamiinid A, K, E ning vees lahustuvad C- ja B-vitamiinid.
Fütohormoonid- kõrge füsioloogilise aktiivsusega madala molekulmassiga ained. Neil on väga madalates kontsentratsioonides reguleeriv toime taimede kasvu- ja arenguprotsessidele. Fütohormoonid jagunevad stimulantideks (tsütokiniinid, auksiinid, giberelliinid) ja inhibiitoriteks (etüleen ja abstsitsiin).
Taimede rakkude, kudede ja elundite elutähtis aktiivsus on tingitud vee olemasolust. Vesi on põhiseaduslik aine. Rakkude ja selle organellide tsütoplasma struktuuri määramisel on see tänu molekulide polaarsusele lahustiks ainevahetuses osalevatele orgaanilistele ja anorgaanilistele ühenditele ning toimib taustkeskkonnana, milles toimuvad kõik biokeemilised protsessid. Kergesti läbi rakkude kestade ja membraanide tungides ringleb vesi vabalt kogu taimes, tagades ainete ülekande ja aidates seeläbi kaasa keha ainevahetusprotsesside ühtsusele. Tänu oma suurele läbipaistvusele ei sega vesi imendumist päikeseenergia klorofüll.
Vee seisund taimerakkudes
Vesi rakus on esitatud mitmel kujul, need on põhimõtteliselt erinevad. Peamised neist on põhi-, solvaat-, kapillaar- ja reservvesi.
Mõned rakku sisenevad veemolekulid moodustavad vesiniksidemeid paljude radikaalmolekulidega orgaaniline aine. Vesiniksidemetega on eriti lihtne moodustada selliseid radikaale:
Seda vee vormi nimetatakse põhiseaduslik . Seda sisaldab rakk, mille tugevus on kuni 90 tuhat barri.
Kuna veemolekulid on dipoolid, moodustavad nad orgaaniliste ainete laetud molekulidega tahkeid agregaate. Sellist vett, mis on elektrilise tõmbejõu abil seotud tsütoplasma orgaaniliste ainete molekulidega, nimetatakse solvaat . Sõltuvalt taimeraku tüübist moodustab solvaadivesi 4–50% selle koguhulgast. Solvaadiveel, nagu ka põhiveel, puudub liikuvus ja see ei ole lahusti.
Suur osa raku veest on kapillaar , kuna see asub makromolekulide vahelistes õõnsustes. Solvaat- ja kapillaarvett hoiab rakk kinni jõuga, mida nimetatakse maatriksipotentsiaaliks. See on võrdne 15-150 baariga.
Reserv nimetatakse vett vakuoolide sees. Vakuoolide sisu on suhkrute, soolade ja mitmete muude ainete lahus. Seetõttu hoiab rakk varuvett jõuga, mille määrab vaakumisisalduse osmootse potentsiaali suurus.
Vee omastamine taimerakkude poolt
Kuna rakkudes puuduvad veemolekulide aktiivsed kandjad, toimub selle liikumine rakkudesse ja rakkudest välja ning naaberrakkude vahel ainult difusiooniseaduste järgi. Seetõttu osutuvad veemolekulide peamiseks tõukejõuks lahustunud aine kontsentratsiooni gradiendid.
Taimerakud, olenevalt nende vanusest ja seisundist, neelavad vett, kasutades järjestikku kolme mehhanismi: imbumine, solvatatsioon ja osmoos.
immitsemine . Kui seemned idanevad, hakkavad need immutusmehhanismi tõttu vett imama. Sel juhul täidetakse protoplasti orgaaniliste ainete vabad vesiniksidemed ja vesi siseneb keskkonnast aktiivselt rakku. Võrreldes teiste rakkudes tegutsevate jõududega on imbumisjõud kolossaalsed. Mõne vesiniksideme puhul ulatuvad need väärtuseni 90 tuhat barri. Samal ajal võivad seemned suhteliselt kuivadel muldadel paisuda ja idaneda. Pärast kõigi vabade vesiniksidemete täitmist imendumine peatub ja aktiveeritakse järgmine veeimamismehhanism.
lahendamine . Solvatatsiooniprotsessis toimub veeimavus, ehitades protoplastide orgaaniliste ainete molekulide ümber hüdratsioonikihte. Raku koguveesisaldus kasvab jätkuvalt. Solvatatsiooni intensiivsus sõltub oluliselt protoplasti keemilisest koostisest. Mida rohkem on rakus hüdrofiilseid aineid, seda täielikumalt kasutatakse solvatatsioonijõude. Hüdrofiilsus väheneb sarjas: valgud -> süsivesikud -> rasvad. Seetõttu imavad valguseemned (herned, oad, oad) massiühiku kohta solvatatsiooni teel suurima koguse vett, tärkliseseemned (nisu, rukis) vahepealse ja õliseemned (lina, päevalill) kõige vähem.
Lahendusjõud on oma võimsuselt väiksemad kui immutusjõud, kuid need on siiski üsna märkimisväärsed ja ulatuvad 100 baarini. Solvatatsiooniprotsessi lõpuks on raku veesisaldus nii suur, et kapillaarniiskus settib ja hakkavad tekkima vakuoolid. Kuid nende moodustumise hetkest solvatatsioon peatub ja vee edasine imendumine on võimalik ainult osmootse mehhanismi tõttu.
Osmoos . Vee omastamise osmootne mehhanism toimib ainult rakkudes, millel on vakuool. Vee liikumise suuna määrab sel juhul osmootsesse süsteemi kuuluvate lahuste osmootsete potentsiaalide suhe.
Rakumahla osmootne potentsiaal, mida tähistatakse R, määratakse järgmise valemiga:
R = iRct,
kus R - rakumahla osmootne potentsiaal
R- gaasikonstant on võrdne 0,0821;
T - temperatuur Kelvini skaalal;
i- isotooniline koefitsient, mis näitab iseloomu elektrolüütiline dissotsiatsioon lahustunud ained.
Isotooniline suhe ise on võrdne
ja= 1 + α ( n + 1),
kus α - elektrolüütilise dissotsiatsiooni aste;
P - ioonide arv, milleks molekul dissotsieerub. Mitteelektrolüütide jaoks P = 1.
Mullalahuse osmootset potentsiaali tähistatakse tavaliselt kreeka tähega π.
Veemolekulid liiguvad alati madalama osmootse potentsiaaliga keskkonnast kõrgema osmootse potentsiaaliga keskkonda. Niisiis, kui rakk on pinnases (välises) lahuses kl R>π, siis vesi siseneb rakkudesse. Vee vool rakku peatub, kui osmootsed potentsiaalid on täielikult võrdsustatud (vaakumimahl lahjendatakse veeimamise sissepääsu juures) või kui rakumembraan jõuab venitatavuse piirini.
Seega saavad rakud keskkonnast vett vaid ühel tingimusel: rakumahla osmootne potentsiaal peab olema suurem kui ümbritseva lahuse osmootne potentsiaal.
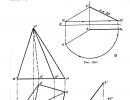
Kui R< π, toimub vee väljavool rakust välislahusesse. Vedelikukao käigus protoplasti maht järk-järgult väheneb, see eemaldub membraanist ja rakku tekivad väikesed õõnsused. Sellist seisundit nimetatakse Plasmolüüs . Plasmolüüsi etapid on näidatud joonisel fig. 3.18.
Kui osmootsete potentsiaalide suhe vastab tingimusele P = π, siis veemolekulide difusiooni ei toimu üldse.
Suur hulk faktilist materjali viitab sellele, et taimede rakumahla osmootne potentsiaal varieerub üsna suurtes piirides. Põllumajandustaimedes juurerakkudes on see tavaliselt amplituudiga 5-10 baari, leherakkudes võib see tõusta kuni 40 baari ja viljarakkudes - kuni 50 baari. Solonchaki taimedes ulatub rakumahla osmootne potentsiaal 100 baarini.

Riis. 3.18.
A - turgori seisundis rakk; B - nurgeline; B - nõgus; G - kumer; D - kramplik; E - kork. 1 - kest; 2 - vakuool; 3 - tsütoplasma; 4 - südamik; 5 - Hecht niidid