Care sunt cele trei tipuri de proteine membranare? Proteine membranare integrale și periferice. Fixare cu o „ancoră” de membrană
Viitorul medicinei este reprezentat de metodele personalizate de influență selectivă asupra sistemelor celulare individuale care sunt responsabile de dezvoltarea și evoluția unei anumite boli. Clasa principală de ținte terapeutice sunt proteinele membranei celulare ca structuri responsabile de asigurarea transmiterii directe a semnalelor în celulă. Deja astăzi, aproape jumătate dintre medicamente acționează în mod specific asupra membranelor celulare și vor exista doar mai multe în viitor. A face cunoștință rol biologic Acest articol este dedicat proteinelor membranare.
Structura și funcția membranei celulare
Din curs şcolar mulți își amintesc dispozitivul unitate structurală organism – celule. Un loc special în structura unei celule vii îl joacă plasmalema (membrană), care separă spațiul intracelular de mediul înconjurător. Astfel, funcția sa principală este de a crea o barieră între conținutul celular și spațiul extracelular. Dar aceasta nu este singura funcție a plasmalemei. Alte funcții ale membranei asociate în principal cu proteinele membranare includ:
- Protectoare (legarea antigenelor și împiedicarea pătrunderii lor în celulă).
- Transport (asigurarea metabolismului între celulă și mediu).
- Semnalizare (complecșii proteici de receptor încorporați asigură iritabilitatea celulelor și răspunsul acesteia la diferite influențe externe).
- Energie - transformarea diferitelor forme de energie: mecanică (flagelele și cilii), electrică (impulsul nervos) și chimică (sinteza moleculelor de acid adenozin trifosforic).
- Contact (oferă comunicare între celule folosind desmozomi și plasmodesmate, precum și pliuri și excrescențe ale plasmalemei).

Structura membranei
Membrana celulară este un strat dublu de lipide. Stratul dublu se formează datorită prezenței în molecula de lipide a două părți cu proprietăți diferite - o regiune hidrofilă și una hidrofobă. Stratul exterior al membranelor este format din „capete” polare cu proprietăți hidrofile, iar „cozile” hidrofobe ale lipidelor sunt direcționate în stratul dublu. Pe lângă lipide, structura membranelor include proteine. În 1972, microbiologii americani S.D. Singer (S. Jonathan Singer) și G.L. Nicholson (Garth L. Nicolson) a propus un model fluid-mozaic al structurii membranei, conform căruia proteinele membranei „plutesc” în stratul dublu lipidic. Acest model a fost extins de biologul german Kai Simons (1997) în ceea ce privește formarea unor anumite regiuni mai dense cu proteine asociate (plute lipidice), care derivă liber în stratul dublu al membranei.
Structura spațială a proteinelor membranare
ÎN celule diferite raportul dintre lipide și proteine este diferit (de la 25 la 75% proteine în ceea ce privește greutatea uscată) și sunt distribuite neuniform. După locație, proteinele pot fi:
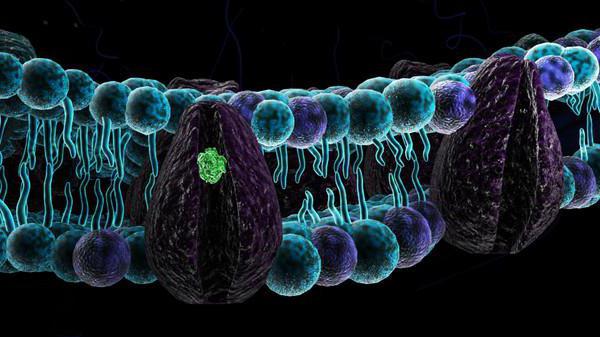
- Integrală (transmembrană) - încorporată în membrană. În același timp, ele pătrund în membrană, uneori în mod repetat. Regiunile lor extracelulare poartă adesea lanțuri de oligozaharide, formând grupuri de glicoproteine.
- Periferic - situat în principal pe partea interioară a membranelor. Comunicarea cu lipidele membranei este asigurată prin legături reversibile de hidrogen.
- Ancorat - situat predominant în exteriorul celulei și „ancora” care le ține la suprafață este o moleculă de lipide scufundată în stratul dublu.

Functionalitate si responsabilitati
Rolul biologic al proteinelor membranare este divers și depinde de structura și localizarea acestora. Printre acestea se numără proteinele receptorului, proteinele canalului (ion și porine), transportori, motoare și clustere structurale de proteine. Toate tipurile de proteine receptorilor membranari, ca răspuns la orice impact, își schimbă structura spațială și formează un răspuns celular. De exemplu, receptorul de insulină reglează fluxul de glucoză în celulă, iar rodopsina în celulele sensibile ale organului vizual declanșează o cascadă de reacții care duc la apariția unui impuls nervos. Rolul proteinelor canalelor membranare este de a transporta ionii și de a menține diferența dintre mediul intern și cel extern. De exemplu, pompele de sodiu-potasiu asigură schimbul de ioni și substanțe corespunzători. Porinele - proteine end-to-end - sunt implicate în transferul moleculelor de apă, transportori - în transferul anumitor substanțe împotriva unui gradient de concentrație. În bacterii și protozoare, mișcarea flagelilor este asigurată de motoarele proteinelor moleculare. Proteinele membranare structurale susțin membrana însăși și asigură interacțiunea altor proteine ale membranei plasmatice.

Proteine pentru membrană, membrană pentru proteine
Membrana este un mediu dinamic și foarte activ, și nu o matrice inertă pentru proteinele care sunt localizate și lucrează în ea. Afectează în mod semnificativ funcționarea proteinelor membranare, iar plutele lipidice, în mișcare, formează noi legături asociative ale moleculelor de proteine. Multe proteine pur și simplu nu funcționează fără parteneri, iar interacțiunea lor intermoleculară este asigurată de natura stratului lipidic al membranelor, a cărui organizare structurală, la rândul său, depinde de proteinele structurale. Tulburările în acest mecanism delicat de interacțiune și interdependență duc la disfuncția proteinelor membranare și la o serie de boli, cum ar fi diabetul și tumorile maligne.
Organizarea structurală
Ideile moderne despre structura și structura proteinelor membranare se bazează pe faptul că, în partea periferică a membranei, cele mai multe dintre ele constau rareori dintr-una, mai adesea din mai multe elice alfa oligomerizante asociate. Mai mult decât atât, tocmai această structură este cheia îndeplinirii funcției. Totuși, clasificarea proteinelor după tipul de structură este cea care poate aduce mult mai multe surprize. Dintre peste o sută de proteine descrise, cea mai studiată proteină membranară după tipul de oligomerizare este glicoforina A (proteina celulelor roșii din sânge). Pentru proteinele transmembranare, situația este mai complicată - a fost descrisă o singură proteină (centrul de reacție fotosintetică al bacteriilor - bacteriorhodopsin). Având în vedere greutatea moleculară mare a proteinelor membranare (10-240 mii daltoni), biologii moleculari au un domeniu larg de cercetare.
Sisteme de semnalizare celulară
Printre toate proteinele membranei plasmatice, un loc special revine proteinelor receptorilor. Ele reglează ce semnale intră în celulă și care nu. În toate organismele pluricelulare și în unele bacterii, informația este transmisă prin molecule speciale (molecule semnal). Printre acești agenți de semnalizare, se numără cei special secretați de celule), formațiuni non-proteice și ioni individuali. Acesta din urmă poate fi eliberat atunci când celulele învecinate sunt deteriorate și declanșează o cascadă de reacții sub formă de durere, principalul mecanism de apărare al organismului.
Ținte pentru farmacologie
Proteinele membranare sunt principalele ținte ale farmacologiei, deoarece acestea sunt punctele prin care trec cele mai multe semnale. „Direcționarea” unui medicament și asigurarea selectivității sale ridicate este sarcina principală atunci când se creează un agent farmacologic. Acțiunea selectivă numai asupra unui anumit tip sau chiar subtip de receptor este un efect asupra unui singur tip de celulă din organism. O astfel de acțiune selectivă poate, de exemplu, să distingă celulele tumorale de celulele normale.

Medicamentele viitorului
Proprietățile și caracteristicile proteinelor membranare sunt deja folosite astăzi în crearea de medicamente de nouă generație. Aceste tehnologii se bazează pe crearea unor structuri farmacologice modulare din mai multe molecule sau nanoparticule „reticulate” între ele. Partea „țintire” recunoaște anumite proteine receptor de pe membrana celulară (de exemplu, cele asociate cu dezvoltarea cancerului). În această parte se adaugă un agent de perturbare a membranei sau un blocant al proceselor de producție de proteine în celulă. Dezvoltarea apoptozei (programul de auto-moarte) sau a unui alt mecanism al cascadei de transformări intracelulare duce la rezultatul dorit al expunerii la un agent farmacologic. Ca urmare, avem un medicament cu un minim de efecte secundare. Primele astfel de medicamente anti-cancer sunt deja în curs de studii clinice și vor deveni în curând cheia unei terapii extrem de eficiente.

Genomica structurală
Știința modernă despre moleculele de proteine se îndreaptă din ce în ce mai mult spre tehnologia de informație. O cale extinsă de cercetare - de a studia și de a descrie tot ceea ce este posibil, de a stoca datele în baze de date computerizate și apoi de a căuta modalități de aplicare a acestor cunoștințe - acesta este scopul biologilor moleculari moderni. Cu doar cincisprezece ani în urmă, proiectul global privind genomul uman a fost lansat și avem deja o hartă secvențială a genelor umane. Al doilea proiect, al cărui scop este de a determina structura spațială a tuturor „proteinelor cheie”, genomica structurală, este încă departe de a fi finalizat. Structura spațială a fost determinată până acum doar pentru 60 de mii din peste cinci milioane de proteine umane. Și chiar dacă oamenii de știință au crescut până acum doar purcei strălucitori și roșii rezistente la frig cu gena somonului, tehnologiile genomice structurale rămân o etapă a cunoștințelor științifice, a cărei aplicare practică nu va întârzia să apară.
CURS
TEMA: „Introducere în histologie. Membrana plasmatica, structura si functiile. Structuri formate de membrana plasmatica"
Histologia în traducere literală- aceasta este știința țesuturilor, dar acest concept nu găzduiește cantitatea cu adevărat mare de material pe care o acoperă această disciplină cu adevărat medicală. Cursul de histologie începe cu studiul citologiei nu atât la nivel luminos-optic, cât la nivel molecular, care medicina modernă intrat logic în etiologia şi patogeneza unui număr de boli. Histologia include, de asemenea, secțiuni separate din cursul embriologiei, nu toate, desigur, ci acea parte a acesteia care abordează problema formării și diferențierii primordiilor tisulare. Și, în sfârșit, histologia este o secțiune mare a histologiei private, adică o secțiune care studiază structura și funcțiile diferitelor organe. Secțiunile enumerate ale cursului de histologie nu lasă nicio îndoială că studiul disciplinei noastre ar trebui efectuat sub aspectul menținerii unității nivelurilor de organizare celulare, tisulare, de organe și de sistem.
Începem histologia prin studierea celulei eucariote, care este cel mai simplu sistem dotat cu viață. Când examinăm o celulă sub un microscop cu lumină, obținem informații despre dimensiunea, forma acesteia, iar această informație este asociată cu prezența limitelor legate de membrană în celule. Odată cu dezvoltarea microscopiei electronice (EM), înțelegerea noastră a membranei ca o linie de divizare clar definită între celulă și mediu schimbat, pentru că s-a dovedit că pe suprafața celulei există structura complexa, constând din următoarele 3 componente:
1. Componentă supramembranară (glicocalice) (5-100 nm)
2. Membrană plasmatică (8-10 nm)
3. Componenta submembranara (zona de variatie a proteinelor citoscheletice)
În același timp, componentele 1 și 3 sunt variabile și depind de tipul de celulă cea mai statică structură pare să fie structura membranei plasmatice, pe care o vom lua în considerare.
Studiul plasmalemei în condiții EM a condus la concluzia că organizarea sa structurală este uniformă, în care are aspectul unei linii trilaminare, unde straturile interior și exterior sunt electrodense, iar stratul mai larg situat între ele pare să să fie transparent la electroni. Acest tip de organizare structurală a membranei indică eterogenitatea sa chimică. Fără a atinge discuția pe această temă, vom prevedea că plasmalema este formată din trei tipuri de substanțe: lipide, proteine și carbohidrați.
Lipidele, care fac parte din membrane, au proprietăți amfifile datorită prezenţei în compoziţia lor atât a grupărilor hidrofile cât şi a celor hidrofobe.
Natura amfipatică a lipidelor membranare promovează formarea unui dublu strat lipidic. În acest caz, în fosfolipidele membranare se disting două domenii: a) fosfat– capul moleculei, proprietăți chimice acest domeniu determină solubilitatea sa în apă și se numește hidrofil
b) lanțuri acil, care sunt acizi grași esterificați – acesta este un domeniu hidrofob.
Tipuri de lipide membranare. 1. Principala clasă de lipide din membranele biologice sunt fosfo(gliceridele) (fosfolipidele), ele formează cadrul
membrana biologică (fig. 1).
Biomembrane- acesta este un strat dublu lipide amfifile(dublu strat lipidic). ÎN mediu acvatic astfel de molecule amfifile formează în mod spontan un dublu strat în care părțile hidrofobe ale moleculelor sunt orientate una spre cealaltă, iar părțile hidrofile către apă (Fig. 2).
Membranele conțin următoarele tipuri de lipide:
1. Fosfolipide
2. Sfingolipide „cap” + 2 „cozi” hidrofobe
3.Glicolipidele
Colesterol (CL)– este situat în membrană în principal în zona mijlocie a stratului dublu, este amfifil și hidrofob(cu excepția unei grupări hidroxi). Compoziția lipidică afectează proprietățile membranelor: raportul proteină/lipide este aproape de 1:1, totuși, tecile de mielină sunt îmbogățite în lipide, iar membranele interne sunt îmbogățite în proteine.
Metode de ambalare pentru lipide amfifile: 1. Bistraturi (membrană lipidică), 2. Lipozomii sunt o veziculă cu două straturi de lipide, în timp ce atât suprafețele interioare cât și cele exterioare sunt polare. 3. Micelele - a treia variantă de organizare a lipidelor amfifile - o veziculă, al cărei perete este format dintr-un strat de lipide, în timp ce capetele lor hidrofobe sunt îndreptate spre centrul micelului și mediul lor intern nu este apos, dar hidrofob.
Cea mai comună formă de ambalare a moleculelor de lipide este formarea lor plat dublu strat membranar. Lipozomii și miceliile sunt forme de transport rapid care asigură transferul de substanțe în și în afara celulei. În medicină, lipozomii sunt folosiți pentru a transporta substanțe solubile în apă, iar miceliile sunt folosite pentru a transporta substanțe solubile în grăsimi.
Proteinele membranare:
1. Integral (inclus în straturile lipidice)
2. Periferic
Integral (proteine transmembranare):
1. Monotopic- (de exemplu, glicoforina. Ei traversează membrana de 1 dată) și sunt receptori, în timp ce exteriorul lor - domeniul extracelular - aparține părții de recunoaștere a moleculei.
2. Politopic– pătrund în mod repetat în membrană – acestea sunt, de asemenea, proteine receptor, dar activează calea de transmitere a semnalului în celulă.
Proteinele membranare asociate cu lipide.
4. Proteinele membranare, asociate cu carbohidrații.
proteine periferice - nu sunt scufundate în stratul dublu lipidic și nu sunt legate covalent de acesta. Ele sunt ținute împreună prin interacțiuni ionice. Proteinele periferice sunt asociate cu proteinele integrale din membrană datorită interacțiunii - proteină-proteină interacțiuni.
Un exemplu de aceste proteine:
1. Spectrină, care se află pe suprafata interioara celule
2. Fibronectină, localizat pe suprafața exterioară a membranei
proteine - constituie de obicei până la 50% din masa membranei. În același timp
proteine integraleîndeplini următoarele funcții:
a) proteinele canalelor ionice
b) proteine receptor
2. Proteine membranare periferice(fibrilare, globulare) îndeplinesc următoarele funcții:
a) extern (proteine receptori și de adeziune)
b) interne – proteine din citoschelet (spectrin, anchirina), proteine ale sistemului mesager secund.
Canale ionice– acestea sunt canale formate din proteine integrale formează un mic por prin care ionii trec de-a lungul unui gradient electrochimic. Cele mai cunoscute canale sunt canalele pentru Na, K, Ca 2, Cl.
Există și canale de apă - acestea sunt acvaporine(eritrocite, rinichi, ochi).
Componentă supramembranară– glicocalix, grosime 50 nm. Acestea sunt regiuni carbohidrate ale glicoproteinelor și glicolipidelor care oferă o sarcină negativă. Sub EM se află un strat liber de densitate moderată care acoperă suprafața exterioară a plasmalemei. Pe langa componentele carbohidrati, glicocalixul contine proteine membranare periferice (semi-integrale). Zonele lor funcționale sunt situate în zona supramembranară - acestea sunt imunoglobuline (Fig. 4).
Funcția glicocalixului: 1. Joacă un rol receptori.
2. Recunoașterea intercelulară.
3. Interacțiuni intercelulare(interacțiuni adezive).
4. R receptorii de histocompatibilitate.
5. Zona de adsorbție a enzimelor(digestia parietala).
6. Receptorii hormonali.
Componenta submembrană sau zona cea mai exterioară a citoplasmei, are de obicei o rigiditate relativă și această zonă este deosebit de bogată în filamente (d 5-10 nm). Se presupune că proteinele integrale care alcătuiesc membrana celulară sunt asociate direct sau indirect cu filamentele de actină situate în zona submembrană. În același timp, s-a dovedit experimental că în timpul agregării proteinelor integrale, actina și miozina situate în această zonă se agregează, de asemenea, ceea ce indică participarea filamentelor de actină la reglarea formei celulei.
Structuri formate de plasmalema
Contururile celulei, chiar și la nivel optic-luminos, nu apar uniforme și netede, iar microscopia electronică a făcut posibilă detectarea și descrierea diferitelor structuri din celulă care reflectă natura specializării sale funcționale. Se disting următoarele structuri:
1. Microvilozități – proeminență a citoplasmei acoperită cu plasmalemă. Citoscheletul microvilus este format dintr-un mănunchi de microfilamente de actină, care sunt țesute în rețeaua terminală a părții apicale a celulelor (Fig. 5). Microvilozitățile unice nu sunt vizibile la nivel optic de lumină. Dacă există un număr semnificativ de ele (până la 2000-3000) în partea apicală a celulei, chiar și cu microscopia luminoasă se distinge o „graniță perie”.
2. Genele – sunt situate în zona apicală a celulei și au două părți (Fig. 6): a) exterior - axonem
B) intern – corp becal
Axonema constă dintr-un complex de microtubuli (9 + 1 perechi) și proteine asociate. Microtubulii sunt formați de proteina tubulină, iar mânerele sunt formate de proteina dineină - aceste proteine împreună formează traductorul chimiomecanic tubulină-dineină.
Corpul bazal este format din 9 tripleți de microtubuli situati la baza ciliului și servește drept matrice pentru organizarea axonemului.
3. Labirint bazal- Acestea sunt invaginări profunde ale plasmalemei bazale cu mitocondrii situate între ele. Acesta este un mecanism de absorbție activă a apei, precum și a ionilor împotriva unui gradient de concentrație.
1. Transport compuși cu greutate moleculară mică realizat în trei moduri:
1. Difuziune simplă
2. Difuzare facilitată
3. Transport activ
Difuziune simplă– compuși organici hidrofobi cu greutate moleculară mică (acizi grași, uree) și molecule neutre (HO, CO, O). Pe măsură ce diferența de concentrație între compartimentele separate de membrană crește, crește și viteza de difuzie.
Difuzare facilitată– substanța trece prin membrană și în direcția gradientului de concentrație, dar cu ajutorul unei proteine de transport – translocaze. Acestea sunt proteine integrale care au specificitate pentru substanțele transportate. Acestea sunt, de exemplu, canalele anionice (eritrocite), canalele K (plasmolema celulelor excitate) și canalele Ca (reticulul sarcoplasmatic). Translocaza pentru H O este acvaporina.
Mecanismul de acțiune al translocazei:
1. Prezența unui canal hidrofil deschis pentru substanțe de o anumită dimensiune și sarcină.
2. Canalul se deschide numai atunci când se leagă un anumit ligand.
3. Nu există un canal ca atare, iar molecula de translocază însăși, care a legat ligandul, se rotește cu 180° în planul membranei.
Transport activ– acesta este transportul folosind aceeași proteină de transport (translocaze), dar împotriva unui gradient de concentraţie. Această mișcare necesită energie.
Spre deosebire de lipide, proteinele de membrană sunt greu de clasificat după structură, este mai potrivit să se subdivizeze principalele tipuri de proteine de membrană în funcție de funcțiile lor. De regulă, proteinele sunt responsabile pentru activitatea funcțională a membranelor. Aceste proteine includ o varietate de enzime, proteine de transport, receptori, proteine canale, proteine care formează pori de apă (acvaporine), precum și diverse proteine structurale și de reglare care asigură diverse funcții ale membranelor celulare. De rol biologic proteinele membranare pot fi împărțite în patru grupe:
1) proteine enzimatice cu activitate catalitică;
2) proteine receptor care leagă în mod specific anumite substanțe;
3) proteine structurale;
4) proteine care asigură interacțiuni intercelulare.
Proteine enzimatice cea mai abundentă dintre toate proteinele membranare. Acestea includ atât proteine integrale (ATPaze membranare care îndeplinesc o funcție de transport) cât și periferice (acetilcolinesteraza, fosfataze acide și alcaline, RNază). Proteinele de transport creează fluxuri stabile anumite substante iar ionii prin membrane. Transportul ionic duce la apariția potențialului transmembranar în toate celulele, precum și la modificările acestuia în celulele nervoase și musculare. Acest din urmă fenomen stă la baza unor proprietăți atât de importante ale acestor țesuturi precum excitabilitatea și conductivitatea.
Proteinele receptorilor sunt proteine care leagă în mod specific anumiți liganzi implicați în transmiterea semnalelor de la o celulă la alta. O astfel de transmisie se realizează în diferite moduri. Receptorii fac adesea parte din complexe membranare mai complexe care conțin proteine călău. De exemplu, în sinapsele nervoase și neuromusculare, molecula de semnalizare (mediatorul) este o anumită substanță cu un nivel molecular scăzut, iar membrana plasmatică conține proteine speciale de receptor conectate la canalele ionice care își schimbă proprietățile atunci când receptorul se leagă de liganzi. Această reacție asigură permeabilitatea membranei pentru diverși ioni (sodiu, potasiu, calciu, clor) și formează potențialul excitant. Unii receptori (de exemplu, receptorul colinergic nicotinic) înșiși sunt canale ionice (datorită includerii de subunități proteice suplimentare în ansamblul receptorului).
În unele cazuri, proteina receptorului nu este un canal ionic, ci este asociată cu o cascadă de semnalizare intracelulară, a cărei activare are loc la legarea unui ligand care transportă informații la receptor. Ca urmare a activării unor astfel de receptori (se numesc metabotropi, spre deosebire de ionotropi, care reglează fluxurile de ioni prin membrană), apare o cascadă. reactii chimice, manageri funcții celulare prin modificari ale metabolismului (de unde si denumirea acestor receptori). Activarea receptorilor metabotropi de către liganzi (aceștia pot fi considerați molecule de semnalizare primare, sau mesageri primari) conduce la producerea de molecule de semnalizare secundare (mesageri secundi) în citoplasma celulei activate.
Proteinele structurale conferă celulelor și organelelor forma lor specifică; oferi unul sau altul proprietăți mecanice(de exemplu, elasticitate) membrană plasmatică; Ele conectează membrana cu citoscheletul, iar în cazul membranei nucleare cu cromozomii. Proteinele membranare structurale, de regulă, nu au proprietăți enzimatice (poate că sunt pur și simplu puțin studiate chimic). Studiul lor este îngreunat în principal de două circumstanțe. În primul rând, proteinele structurale sunt „tăcute” – nu au activitate enzimatică cunoscută. În al doilea rând, proteinele structurale au regiuni hidrofobe extinse în moleculele lor. Când sunt purificate, formează asociații apropiate între ele sau cu lipide, ceea ce le complică studiul.
Proteina neurospecifică B-50 este una dintre principalele proteine structurale fosforilate ale membranelor plasmatice ale contactelor sinaptice. Prin metode imunochimice s-a stabilit că este localizat predominant în membranele presinaptice. Greutate moleculară proteine 48 kPa. Este un substrat endogen al protein kinazei C dependente de protein kinazei C. Activatorii proteinei kinazei C stimulează procesul de transmitere sinaptică în feliile hipocampice. Fosforilarea proteinei B-50 duce la o creștere a timpului stării excitate a sinapsei, ceea ce ajută la menținerea canalelor ionice în starea activată (deschisă) (în unele publicații acest fenomen este numit starea sinapsei). Efectul proteinei fosforilate B-50 asupra metabolismului fosfoinozitidelor poate fi unul dintre motivele acestui fenomen. Interesant este că pe măsură ce corpul îmbătrânește, intensitatea fosforilării proteinei B-50 din creier scade, ceea ce poate determina o scădere a plasticității sinapselor.
Alte dovezi ale rolului fosforilării proteinei B-50 în funcționarea sinapselor au fost obținute în experimente in vitro, care au confirmat că neuropeptida - un fragment de ACTH1_24 - este de 10 ori mai eficientă în inhibarea fosforilării B-50 în membranele sinaptice din regiunea septală a creierului decât în membranele întregului creier
Grupul proteinelor membranare include și multe proteine enzimatice care formează canale ionice - Na/K- și Ca-ATPaze, proteine receptor, sinapsine etc.
Plasmină- serin proteinaza, în plasma sanguină acționează în principal ca o enzimă trombolitică și, de asemenea, degradează multe componente ale matricei extracelulare. În creier, plasmina este implicată în numeroase funcții precum plasticitatea neuronală, învățarea și memoria. Activarea sistemului plasminogen este observată în creier în timpul și în primele zile după un accident vascular cerebral. În boala Alzheimer, dimpotrivă, există o scădere a nivelului de plasmină din țesutul cerebral.
Plasmina, care face parte din membranele plasmatice ale neuronilor, este în asociere cu plute bogate în colesterol, care sunt considerate a fi locul formării predominante a beta-A amiloidului. Aceasta indică o legătură funcțională între plasmină, colesterol și metabolismul creierului.
Enzima de conversie a endotepinei(ECE-1) este o altă enzimă de degradare a amiloidului, care este 37% omoloagă cu NEP în secvența de aminoacizi ECE-1 este, de asemenea, o metaloprotează dependentă de zinc legată de membrană, la fel ca NEP, este capabilă să se degradeze. număr mare biologic substanțe active, inclusiv bradikinină, neurotensină, angiotensină-1 și lanțul B de insulină. Spre deosebire de NEP, ECE-1 există sub formă de dimeri, ale căror subunități sunt conectate printr-o legătură disulfurică.
Aceste două metaloproteaze (ECE-1 și NEP) diferă și ca sensibilitate la inhibitori. Inhibarea NEP necesită concentrații nanomolare de tiorfan și fosforamidonă, în timp ce ECE-1 este inhibată de concentrațiile micromolare de fosforamidonă și nu este sensibilă la tiorfan.
ECE-1 se găsește în multe organe și țesuturi. Celulele endoteliale sunt cele mai îmbogățite cu această enzimă în care este exprimată țesut nervos si muschii. În celulele musculare netede, ECE-1 se găsește în complex cu filamente de alfa-actină.
În prezent, sunt cunoscuți patru izofori ECE-1 umani (1a, 1b, 1c și 1d), care nu au diferențe catalitice semnificative, dar diferă în localizarea subcelulară. Izoformele 1a, 1b, 1c și 1d se găsesc pe suprafața celulei, în timp ce ECE-1 este o formă intracelulară localizată în aparatul Golgi.
A fost descoperită o altă proteină asemănătoare cu ECE-1, care este localizată în principal în creier și în cantități mici în celulele endoteliale și musculare netede. Structura sa primară este 59°/o identică cu secvența de aminoacizi a ECE-1. Este desemnat ca ECE-2 și diferă de ECE-1 prin faptul că are un pH optim mai acid.
| | | | | | | | | 10 |
De regulă, proteinele sunt responsabile pentru activitatea funcțională a membranelor. Acestea includ o varietate de enzime, proteine de transport, receptori, canale, pori etc. Anterior, se credea că proteinele membranare au structură secundară exclusiv a proteinei pliate β, dar aceste studii au arătat că membranele conțin un număr mare de elice α. Studiile ulterioare au arătat că proteinele membranare pot pătrunde adânc în stratul dublu lipidic sau chiar pot pătrunde în el, iar stabilizarea lor se realizează datorită hidrofobei...
Distribuiți-vă munca pe rețelele sociale
Dacă această lucrare nu vă convine, în partea de jos a paginii există o listă cu lucrări similare. De asemenea, puteți utiliza butonul de căutare
Cursul 5
Structura și funcțiile proteinelor membranare
Membranele celulare conțin proteine de la 20 la 80% (în greutate). De regulă, proteinele sunt responsabile pentru activitatea funcțională a membranelor. Acestea includ o varietate de enzime, proteine de transport, receptori, canale, pori etc. etc., care asigură funcțiile unice ale fiecărei membrane. Primele progrese în studiul proteinelor membranare au fost realizate atunci când biochimiștii au învățat să folosească detergenți pentru a izola proteinele membranei într-o formă activă funcțional. Acestea au fost lucrări privind studiul complexelor enzimatice ale membranei interioare a mitocondriilor. Anterior, se credea că proteinele membranare au o structură pliată exclusiv β (structură secundară a proteinei), dar aceste studii au arătat că membranele conțin un număr mare de elice α. Helixul β este mult mai puțin comun, dar i se acordă totuși un aspect important semnificație biologică. Cert este că în zonele înconjurate de lipide, helixul β este un cilindru gol, în peretele exterior al căruia sunt concentrate reziduuri de aminoacizi nepolare (hidrofobe), iar în peretele interior sunt hidrofile. Un astfel de cilindru ar putea forma un canal în membrană prin care trec liber ionii și substanțele solubile în apă. Studiile ulterioare au arătat că proteinele membranare pot pătrunde adânc în stratul dublu lipidic sau chiar pot pătrunde și sunt stabilizate prin interacțiuni hidrofobe. Există cel puțin patru tipuri de aranjare a proteinelor în membrane: Primul tip este transmembranar, când proteina pătrunde în întreaga membrană, iar regiunea hidrofobă a proteinei are o configurație α. Molecula de bacteriorhodopsină din Halobacterium halobium elicele sale α traversează secvenţial stratul dublu; Al doilea tip este legarea folosind o ancoră hidrofobă, când proteina are o regiune scurtă constând din resturi de aminoacizi hidrofobe în apropierea capătului carboxil. Aceasta este așa-numita ancora hidrofobă, care poate fi îndepărtată prin proteoliză, iar proteina eliberată devine solubilă în apă. Acest aranjament în membrană este caracteristic multor citocromi. Al treilea tip este legarea de suprafața stratului dublu, atunci când interacțiunea proteinelor este în principal de natură electrostatică sau hidrofobă. Acest tip de interacțiune poate fi folosit ca o completare la alte interacțiuni, cum ar fi ancorarea transmembranară. Al patrulea tip se leagă de proteinele încorporate în stratul dublu, care este atunci când unele proteine se leagă de proteinele care sunt situate în interiorul stratului dublu lipidic. De exemplu, F1 - partea H+ - ATPaza care se leagă de F 0 partea scufundată în membrană, precum și unele proteine citoscheletice.
În miez idei moderne Structura proteinelor membranare se bazează pe ideea că lanțul lor polipeptidic este pliat astfel încât să formeze o suprafață nepolară, hidrofobă în contact cu regiunea nepolară a stratului dublu lipidic. Domeniile polare ale unei molecule de proteine pot interacționa cu capetele polare ale lipidelor de pe suprafața stratului dublu. Multe proteine sunt transmembranare și se întind pe stratul dublu. Unele proteine par să fie asociate cu membrana doar prin interacțiunea lor cu alte proteine.
Multe proteine membranare se asociază de obicei cu membrana prin interacțiuni necovalente. Cu toate acestea, există proteine care sunt legate covalent de lipide. Multe proteine ale membranei plasmatice aparțin clasei de glicoproteine. Reziduurile de carbohidrați ale acestor proteine sunt întotdeauna localizate în exteriorul membranei plasmatice.
De obicei, proteinele membranare sunt împărțite în externe (periferice) și interne (integrale). În acest caz, criteriul este gradul de severitate al procesării necesare pentru extragerea acestor proteine din membrană. Proteinele periferice sunt eliberate atunci când membranele sunt spălate cu soluții tampon cu tărie ionică scăzută, pH scăzut sau, dimpotrivă, ridicat și în prezența agenților de chelare (de exemplu, EDTA) care leagă cationii divalenți. Este adesea foarte dificil să distingem proteinele membranei periferice de proteinele legate de membrană în timpul procesului de eliberare.
Pentru a elibera proteinele membranare integrale, este necesar să folosiți detergenți sau chiar solvenți organici.
Multe proteine membranare eucariote și procariote sunt legate covalent de lipide, care sunt adăugate la polipeptidă după translație.
Proteinele membranei asociate covalent cu lipide
(O) Proteine de care este atașat acidul miristic
(B) Proteine de care este atașat acidul palmitic
(ÎN) Proteine cu ancora de glicozilfosfatidilinozitol
4. Moleculă de adeziune a celulelor nervoase |
În unele cazuri, aceste lipide joacă rolul unei ancora hidrofobe, cu ajutorul căreia proteina este atașată de membrană. În alte cazuri, lipidele sunt susceptibile de a servi ca ajutor în migrarea proteinelor în regiunea celulară corespunzătoare sau (ca în cazul proteinelor învelișului viral) în fuziunea membranei.
La procariote, proteina cea mai complet caracterizată este lipoproteina lui Brown, principala lipoproteină a membranei exterioare. E. coli . Forma matură a acestei proteine conține acilglicerol, care este legat printr-o legătură tioester de N cisteină terminală. In plus, N Aminoacidul terminal este legat de acidul gras printr-o legătură amidă. Forma de penicilază legată de membrană este atașată de membrana citoplasmatică prin N acilglicerol terminal similar cu lipoproteinele membranare.
Proteinele de membrană ale eucariotelor sunt asociate covalent cu lipide, așa cum se arată în tabel, ele pot fi împărțite în trei clase. Proteinele din primele două clase, aparent, sunt localizate în principal pe suprafața citoplasmatică a membranei plasmatice, iar proteinele din clasa a treia pe suprafața exterioară.
Există proteine de membrană care sunt legate covalent de carbohidrați. Acestea includ proteine de suprafață celulară care îndeplinesc în principal funcțiile de transport și recepție. Încă nu este clar ce se întâmplă aici. Acest lucru se poate datora faptului că proteinele trebuie sortate atunci când sunt direcționate către membrana plasmatică. Reziduurile de zahăr pot proteja proteina de proteoliză sau pot participa la recunoaștere sau aderență. Prin urmare, reziduurile de zahăr din glicoproteinele membranei sunt localizate exclusiv pe partea exterioară a membranei.
Se pot distinge două clase principale de structuri oligozaharide ale glicoproteinelor membranare: 1) N oligozaharide glicozidice legate de proteine prin grupul amidic al asparginei; 2) Oligozaharide O-glicozidice legate prin grupări hidroxil serină și treonină. Această clasă de oligozaharide este formată din trei subclase.
- Un complex simplu sau bogat în manoză în care oligozaharida conține manoză și N acetilglucozamină.
- Un complex normal în care miezul bogat în manoză are ramuri laterale suplimentare care conțin alte resturi de zaharide, cum ar fi acidul sialic.
- Complex mare care este asociat cu transportorul anionic al membranei eritrocitare
Majoritatea oligozaharidelor glicoproteice membranare aparțin subclasei 1 sau 2.
Proteinele membranare ale bacteriilor
După cum sa menționat mai sus, proteinele din membrana citoplasmatică reprezintă aproximativ 50% din suprafața acesteia. Aproximativ 10% din membrană este formată din complexe proteină-lipidă strâns legate. Molecula oricărei proteine încorporate în membrană este înconjurată de 45 × 130 sau mai multe molecule de lipide. Aproximativ jumătate din lipidele libere sunt asociate cu proteinele membranei periferice.
Compoziția proteică a membranei citoplasmatice a bacteriilor este mai diversă decât compoziția lipidică. Astfel, în membrana citoplasmatică E. coli K 12 Au fost descoperite aproximativ 120 de proteine diferite. În funcție de orientarea membranei și de natura conexiunii cu stratul dublu lipidic, așa cum sa menționat mai sus, proteinele sunt împărțite în integrale și periferice. Proteinele bacteriene periferice includ o serie de enzime cum ar fi NADH dehidrogenaza, malat dehidrogenaza etc., precum și unele proteine care fac parte din complexul ATPază. Acest complex este un grup de subunități proteice dispuse într-un anumit mod, în contact cu citoplasma, spațiul periplasmatic și formând un canal în membrana prin care trece protonul. Zona complexului desemnată F 1 , cufundat în citoplasmă, și și cu componente ale site-ului F 0 Laturile hidrofobe ale moleculelor sunt scufundate în membrană. Subunitate b parțial scufundat în membrană cu partea sa hidrofobă și comunică membrana și părțile citoplasmatice ale complexului enzimatic, precum și legătura sintezei ATP în regiune F 1 cu potenţial de protoni în membrană. subunitățile a, b și c furnizează un canal de protoni. Alte componente ale complexului îi asigură integritatea structurală și funcțională.
Spre proteine integrale E. coli, care necesită lipide pentru manifestarea activității enzimatice, includ succinat dehidrogenaza, citocromul b . Antibioticele gramicidina A, alameticina, amfotericina si nistacina au proprietati foarte interesante. Când interacționează cu membrana bacteriană, ele devin proteine integrale (antibioticele sunt polipeptide și macrocicluri).
Gramicidina A este o peptidă hidrofobă constând din 15 L-D -aminoacizi. Când este înglobat într-o membrană, formează canale care permit cationii monovalenți să treacă prin. Acest canal, care formează gramicidina A, a fost cel mai complet caracterizat. Canalul este format din două molecule de gramicidină A. Ca urmare a alternanței L- și D - aminoacizii formează o spirală în care lanțurile laterale sunt situate în exterior, iar grupările carboxil ale scheletului sunt în interiorul canalului. Acest tip de helix nu se găsește în nicio altă proteină și se formează din alternanța neobișnuită a stereoizomerilor aminoacizilor din gramicidina A. Canalul gramicidinei, așa cum sa menționat mai sus, este selectiv pentru cationi. Prin ea trec mici cationi anorganici și organici, în același timp permeabilitate Cl - este egal cu zero.
Alameticina este un antibiotic peptidic cu 20 de reziduuri de aminoacizi, capabil să formeze canale excitabile electric în membrană. Secvența de aminoacizi a alameticinei include resturile neobișnuite de acid aminobutiric și L fenilalanina. Când este legat de o membrană, spre deosebire de gramicidina A, formează un por. Este mult mai mic ca dimensiune decât canalul care formează gramicidina A. Acest lucru se datorează în primul rând faptului că spațiul din jurul helixului α este prea mic pentru a trece un ion.
Antibioticele marcolide precum nistatina și amfotericina se leagă de colesterol și formează canale. Canalele formează 8 × 10 molecule din aceste antibiotice poliene, prin care, totuși, ionii pătrund la viteze scăzute.
Alte lucrări similare care vă pot interesa.vshm> |
|||
| 21572. | STRUCTURA ŞI FUNCŢIA PROTEINELOR | 227,74 KB | |
| Conținutul de proteine din corpul uman este mai mare decât conținutul de lipide al carbohidraților. Predominanța proteinelor în țesuturi în comparație cu alte substanțe este relevată la calcularea conținutului de proteine pe masa uscată de țesuturi. Conținutul de proteine din diferite țesuturi fluctuează într-un anumit interval. | |||
| 17723. | Cerebel, structură și funcții | 22,22 KB | |
| 3 Structura generală creier. ÎN sistemul nervos De asemenea, disting partea centrală a sistemului nervos central, care este reprezentată de creier și măduva spinării, și partea periferică, care include nervi, celule nervoase, noduri nervoase, ganglioni și plexuri, care se află topografic în afara măduvei spinării și a creierului. . Obiectul de studiu este anatomia creierului. Acest subiect și obiect implică formularea și rezolvarea următoarelor sarcini: descrieți planul general al structurii creierului evidențiați structura anatomică; | |||
| 5955. | Organele plantelor: funcțiile, structura și metamorfozele lor. | 16,94 KB | |
| Organele florale sunt frunze modificate: frunzele tegumentare formează sepale și petale, iar frunzele care formează spori dau naștere la stamine și pistil. Lăstarul include: o tulpină b frunze c muguri vegetativi d flori e fructe. O tulpină este un organ vegetativ al unei plante care îndeplinește numeroase funcții: poartă frunze sau o coroană grea de ramuri și frunze; leagă rădăcinile și frunzele; pe el se formează flori; apa si mineralele se deplaseaza de-a lungul ei compuși organici; tulpini tinere... | |||
| 5067. | Mușchii netezi. Structură, funcții, mecanism de contracție | 134,79 KB | |
| Mușchi sau mușchi din lat. Mușchii vă permit să mișcați părți ale corpului și să exprimați gândurile și sentimentele în acțiuni. Mușchii netezi fac parte din unele organele interneși să participe la asigurarea funcțiilor îndeplinite de aceste organisme. | |||
| 6233. | Structura și funcțiile nucleului. Morfologia și compoziția chimică a nucleului | 10,22 KB | |
| Nucleii sunt de obicei separați de citoplasmă printr-o limită clară. Bacteriile și algele albastru-verzi nu au un nucleu format: nucleul lor nu are un nucleol și nu este separat de citoplasmă printr-o membrană nucleară clar definită și se numește nucleoid. Forma miezului. | |||
| 9495. | Clasificare, caracteristici ale gamei de materii prime din blană și semifabricate din blană, structura pielii de blană, structura părului și varietatea formelor sale, tehnologia de fabricare a blănurilor | 1,05 MB | |
| Plăcile de blană sunt benzi de o anumită formă, cusute din piei îmbrăcate selectate și destinate tăierii în părți ale produselor din blană. Tipurile de materii prime pentru blană de iarnă includ piei și piei de animale purtătoare de blană, a căror extracție se realizează în principal în ora de iarna când calitatea pieilor este deosebit de ridicată. STRUCTURA ȘI COMPOZIȚIA CHIMICĂ A BLĂNIILOR ȘI A PIELELOR DE BLANĂ DE ȘHEPPENS MATERIEI PRIME CONCEPTUL DE TOPOGRAFIA PIELELOR Pielea este învelișul exterior al unui animal, separat de carcasa acestuia și format din țesut cutanat și păr. tu... | |||
| 8011. | Proprietățile lipidelor membranare | 10,13 KB | |
| Unele lipide ajută la stabilizarea zonelor foarte curbate ale membranei, formează contact între membrane sau leagă anumite proteine, deoarece forma acestor molecule favorizează împachetarea dorită a stratului dublu în zonele corespunzătoare ale membranei. Starea lichidă este înțeleasă ca capacitatea moleculelor de fosfolipide de a se roti și de a se mișca lateral în lobul corespondent al membranei. Sunt alungite și orientate perpendicular pe planul membranei. În starea de cristal lichid, moleculele de acizi grași sunt mobile, dar... | |||
| 8014. | Compoziția chimică a lipidelor membranare | 10,81 KB | |
| În primul rând, acest lucru se datorează numeroaselor funcții pe care lipidele le îndeplinesc în membrane. Acidul fosfatidic se găsește sub formă liberă în membranele bacteriene în cantități mici, de obicei cu reziduuri de alcooli aminoacizi și altele atașate. Aceste lipide sunt esteri de acizi grași și glicerol și sunt larg reprezentate în multe membrane ale celulelor eucariote și procariote. cu excepția arheobacteriilor. Se găsesc în cantități mari în membrana interioară a mitocondriilor, în membrana cloroplastelor și în unele bacterii... | |||
| 21479. | METABOLISMUL PROTEINELOR | 150,03 KB | |
| Există trei tipuri de bilanţ de azot: bilanţ de azot bilanţ de azot pozitiv bilanţ de azot negativ Cu un bilanţ de azot pozitiv, aportul de azot prevalează asupra eliberării acestuia. În cazul bolilor de rinichi, este posibil un bilanț fals pozitiv de azot, în care produsele finale ale metabolismului azotului sunt reținute în organism. Cu un bilanţ negativ de azot, excreţia de azot predomină asupra aportului său. Această afecțiune este posibilă cu boli precum tuberculoza, reumatismul, oncologia... | |||
| 15073. | Luarea în considerare a electrozilor membranari (ion-selectivi) cu diferite tipuri de membrane | 127,48 KB | |
| În acest scop, există diverși electrozi ion-selectivi, a căror caracteristică principală este așa-numita selectivitate la un anumit tip de ion. Electrozii cu membrane lichide și peliculoase Membranele lichide sunt soluții în solvenți organici ale schimbătoarelor de ioni, schimbătoarelor de cationi lichide sau schimbătoarelor de anioni sau chelaților neutri separați de solutii apoase Pereți despărțitori din sticlă polimer poroasă neutră sau altele. În prezent, industria produce electrozi cu film ion-selectiv pentru cationi N K NH4 Ca2... | |||
Chimie biologică Lelevich Vladimir Valeryanovich
Proteinele membranare.
Proteinele membranare.
Proteinele membranare sunt responsabile pentru activitatea funcțională a membranelor și reprezintă 30 până la 70%. Proteinele membranei diferă în funcție de poziția lor în membrană. Ele pot pătrunde adânc în stratul dublu lipidic sau chiar îl pot pătrunde - proteine integrale, în moduri diferite se atașează la membrană - proteine de suprafață, sau contact covalent cu aceasta - proteine ancorate. Proteinele de suprafață sunt aproape întotdeauna glicozilate. Reziduurile de oligozaharide protejează proteina de proteoliză și sunt implicate în recunoașterea și aderența ligandului.
Proteinele localizate în membrană îndeplinesc funcții structurale și specifice:
1. transport;
2. enzimatic;
3. receptor;
4. antigenic.
Din cartea Fă pe oricine, dar NU un crocodil! de Orsag MihaiDar veverițele? În anii șaizeci, am încercat în mod repetat să am veverițe în casă, dar fiecare astfel de încercare s-a încheiat în cel mai trist mod. După ceva timp, veverițele s-au slăbit, membrele posterioare au fost luate și nefericite animale au murit în convulsii. La început eu
Din cartea The Human Genome: An Encyclopedia Written in Four Letters autor Din cartea Genomul uman [Enciclopedia scrisă în patru litere] autor Tarantul Viaceslav ZalmanovichNu toate genele codifică proteine În primul rând, trebuie remarcat faptul că, pe lângă genele care codifică proteine, genomul conține și gene care sintetizează ARN, care nu sunt ARNm (adică nu codifică proteine), dar efectuează o serie de activități independente. funcții importante în celule. ÎN
Din cartea Ceas viu de Ward Ritchie11. Veverițe în roată Pentru a ne familiariza cu căutarea modernă a unei soluții la problema ceasurilor vii, să ne întoarcem la cercetările efectuate de biologi în ultimii ani. Printre primele, poate, este opera Patriciei de Courcy În 1955, de Courcy a primit o diplomă
Din cartea Biologie [ Ghid complet pentru a se pregăti pentru examenul de stat unificat] autor Lerner Georgy Isaakovich Din cartea Blood: River of Life [De la legendele antice la descoperiri științifice] de Isaac AsimovCapitolul 11 Aceste proteine mobile La începutul capitolului precedent, am menționat că componentele organice ale alimentelor sunt împărțite în trei grupe. Am vorbit despre una dintre aceste grupe: carbohidrații. În continuare, ar fi logic să trecem la proteine, deoarece metabolismul lor în organism are loc în paralel cu
Din cartea Logic of Case [Despre natură și origine evolutie biologica] autor Kunin Evgheniei ViktoroviciCapitolul 12 Originea vieții. Apariția translației, replicării, metabolismului și membranelor: abordări biologice, geochimice și cosmologice Trans. A. Neizvestny În capitolul precedent am discutat posibile scenarii pentru apariția celulelor și (sperăm că) am realizat
Din cartea Genele și dezvoltarea corpului autor Neyfakh Alexander Alexandrovici2. Proteinele cromatinei Știm deja că cromatina constă din ADN și histone în cantități egale de greutate și proteine non-histone (HBP), dintre care în regiunile inactive ale cromozomului există doar 0,2 greutăți ADN, iar în regiunile active - mai mult de 1,2. (în medie, NGB este mai mic decât ADN-ul). Știm și că histones
Din cartea Animal World autor Sitnikov Vitali Pavlovici Din carte Starea actuală biosferei și politicii de mediu autorul Kolesnik Yu.4.1. Formarea membranelor este baza pentru începutul vieții Având în vedere diversitatea gigantică a organismelor vii moderne, putem ajunge la concluzia că au existat multe căi de dezvoltare provenite din forme de viață relicte. De fapt, studiul evoluției moleculare
Din cartea Secretele eredității umane autor Afonkin Serghei IurieviciCelule, proteine și gene Viața este un mod de existență a corpurilor proteice. F. Engels Corpul nostru este un imperiu de celule, fiecare dintre acestea fiind o fabrică în miniatură pentru producerea de proteine. Multe dintre aceste macromolecule esențiale pot fi excretate din organism în
Din cartea Antropologie și concepte de biologie autor Kurchanov Nikolai AnatolieviciProteine Proteinele sunt de o importanță capitală în viața organismelor. Diversitatea enormă a ființelor vii este în mare măsură determinată de diferențele în compoziția proteinelor prezente în corpurile lor. De exemplu, mai mult de 5 milioane dintre ele sunt cunoscute în corpul uman. Proteinele sunt polimeri.
Din cartea Chimie biologică autor Lelevici Vladimir ValerianoviciCompoziția chimică membranelor Membranele sunt compuse din molecule de lipide și proteine, ale căror cantități relative variază mult între diferitele membrane. Carbohidrații sunt conținuti sub formă de glicoproteine, glicolipide și alcătuiesc 0,5%-10% din substanțele membranei. Conform mozaicului fluid
Din cartea autoruluiLipidele membranare. Lipidele membranare sunt molecule amfifile, adică. molecula conține atât grupări hidrofile (capete polare), cât și radicali alifatici (cozi hidrofobe), care formează spontan un strat dublu în care cozile lipidelor se confruntă una cu cealaltă. Grosime
Din cartea autoruluiProteine Valoarea nutritivă a proteinelor este asigurată de prezența aminoacizilor esențiali, ale căror schelete de hidrocarburi nu pot fi sintetizate în corpul uman și, în consecință, trebuie aprovizionate cu alimente. Ele sunt, de asemenea, principalele surse de azot. Alocatie zilnica
Din cartea autoruluiProteinele din țesutul muscular Există trei grupe de proteine: 1. proteine miofibrilare – 45%;2. proteine sarcoplasmatice – 35%;3. proteine stromale – 20%. Acest grup include: 1. miozina; 2. actină;3. actomiozină, precum și așa-numitele proteine de reglare: 4. tropomiozina;5.





