Who formulated the law of germinal similarity. Individual and historical development. Law of germinal similarity. Biogenetic law. Recapitulation. Biogenetic law and embryonic variability
Ontogenesis – implementation of genetic information occurring at all stages.
Ontogenesis is a genetically controlled process. During ontogenesis, the genotype is realized and the phenotype is formed.
Ontogenesis is the individual development of an organism, a set of successive morphological, physiological and biochemical transformations undergone by the organism from the moment of its inception to the end of life. O. includes growth, that is, an increase in body weight, its size, and differentiation. The term "O." introduced by E. Haeckel (1866) when he formulated the biogenetic law.
The first attempt at a historical substantiation of O. was made by I. f. Meckel. The problem of the relationship between O. and phylogeny was posed by C. Darwin and developed by F. Müller, E. Haeckel and others. All new evolutionary traits associated with changes in heredity arise in O., but only those that contribute to better adaptation of the organism to living conditions are preserved in the process of natural selection and transmitted to subsequent generations, that is, they are fixed in evolution. Knowledge of the laws, causes, and factors of oxygen serves as a scientific basis for finding means of influencing the development of plants, animals, and humans, which is of utmost importance for the practice of crop and livestock production, as well as for medicine.
Phylogeny is the historical development of organisms. The term was introduced in German. evolutionist E. Haeckel in 1866. The main task in the study of physiology is to reconstruct the evolutionary transformations of animals, plants, and microorganisms, and to establish on this basis their origin and related relationships between the taxa to which the studied organisms belong. For this purpose, E. Haeckel developed the method of “triple parallelism,” which allows, by comparing data from three sciences—morphology, embryology and paleontology—to reconstruct the course of historical development of the systematic group being studied.
Law of germinal similarity
Explorers of the early 19th century. for the first time they began to pay attention to the similarity of the stages of development of embryos of higher animals with the stages of complexity of organization leading from lowly organized forms to progressive ones. Comparing the stages of development of embryos of different species and classes of chordates, K. Baer made the following conclusions.
1. Embryos of animals of the same type are similar in the early stages of development.
2. They consistently move in their development from more general characteristics of the type to more and more specific ones. Lastly, signs develop that indicate that the embryo belongs to a certain genus, species, and, finally, individual traits.
3. Embryos of different representatives of the same type gradually become separated from each other.
K. Baer, not being an evolutionist, could not connect the patterns of individual development he discovered with the process of phylogenesis. Therefore, the generalizations he made had the meaning of no more than empirical rules.
The subsequent development of the evolutionary idea made it possible to explain the similarity of early embryos by their historical kinship, and their acquisition of more and more specific features with gradual isolation from each other - the actual isolation of the corresponding classes, orders, families, genera and species in the process of evolution.
Soon after the discovery of the law of embryonic similarity, Charles Darwin showed that this law testifies to the common origin and unity of the initial stages of evolution within the type.
Biogenetic law Haeckel-Müller: each living creature in its individual development (ontogenesis) repeats to a certain extent the forms passed by its ancestors or its species (phylogeny).
Ontogenesis - repetition of phylogeny
Comparing the ontogenesis of crustaceans with the morphology of their extinct ancestors, F. Muller concluded that living crustaceans in their development repeat the path traversed by their ancestors. The transformation of ontogeny in evolution, according to F. Muller, is carried out due to its lengthening by adding additional stages or extensions to it. Based on these observations, as well as studying the development of chordates, E. Haeckel (1866) formulated the basic biogenetic law, according to which Ontogenesis is a short and rapid repetition of phylogeny.
The repetition of structures characteristic of ancestors in the embryogenesis of descendants is called recapitulations. Recapitulate not only morphological characters - notochord, gill slits and gill arches in all chordates, but also features of biochemical organization and physiology. Thus, in the evolution of vertebrates, there is a gradual loss of enzymes necessary for the breakdown of uric acid, a product of purine metabolism. In most invertebrates, the final product of the breakdown of uric acid is ammonia, in amphibians and fish it is urea, in many reptiles it is allantoin, and in some mammals uric acid is not broken down at all and is excreted in the urine. In the embryogenesis of mammals and humans, biochemical and physiological recapitulations are noted: the release of ammonia by early embryos, later urea, then allantoin, and in the last stages of development - uric acid.
However, in the ontogenesis of highly organized organisms, a strict repetition of the stages of historical development is not always observed, as follows from the biogenetic law. Thus, the human embryo never repeats the adult stages of fish, amphibians, reptiles and mammals, but is similar in a number of features only to their embryos. The early stages of development remain the most conservative, due to which they recapitulate more completely than the later ones. This is due to the fact that one of the most important mechanisms for integrating the early stages of embryogenesis is embryonic induction, and the structures of the embryo that form first, such as the notochord, neural tube, pharynx, gut and somites, represent the organizational centers of the embryo, from which depends on the entire course of development.
The genetic basis of recapitulation lies in the unity of the mechanisms of genetic control of development, which is preserved on the basis of common genes for the regulation of ontogenesis, which are inherited by related groups of organisms from common ancestors.
Recapitulation(from Latin recapitulatio - repetition) - a concept used in biology to denote the repetition in individual development of characteristics characteristic of an earlier stage of evolutionary development.
Ticket 96.
Ontogenesis as the basis of phylogeny. Cenogenesis. Autonomy of ontogeny. Philembryogenesis. The doctrine of A.N. Severtsov about phylembryogenesis. Mechanisms of their occurrence. Heterochronies and heterotopies of biological structures in the evolution of ontogenesis.
Relying only on the basic biogenetic law, it is impossible to explain the process of evolution: the endless repetition of what has been passed does not in itself give birth to something new. Since life exists on Earth thanks to the change of generations of specific organisms, its evolution occurs due to changes occurring in their ontogenies. These changes boil down to the fact that specific ontogenies deviate from the path laid out by ancestral forms and acquire new features.
Such deviations include, for example, cenogenesis - adaptations that arise in embryos or larvae and adapt them to the characteristics of their environment. In adult organisms, cenogenesis is not preserved. Examples of coenogenesis are horny formations in the mouth of the larvae of tailless amphibians, which make it easier for them to feed on plant foods. During the process of metamorphosis in the frog, they disappear and the digestive system is rebuilt to feed on insects and worms. Coenogenesis in amniotes includes the embryonic membranes, yolk sac and allantois, and in placental mammals and humans they also include the placenta and umbilical cord.
Cenogenesis, appearing only in the early stages of ontogenesis, does not change the type of organization of the adult organism, but provides a higher probability of survival of the offspring. They may be accompanied by a decrease in fertility and a lengthening of the embryonic or larval period, due to which the organism in the postembryonic or postlarval period of development turns out to be more mature and active. Having arisen and proven useful, cenogeneses will be reproduced in subsequent generations. Thus, the amnion, which first appeared in the ancestors of reptiles in the Carboniferous period of the Paleozoic era, is reproduced in all vertebrates developing on land, both in oviparous reptiles and birds, and in placental mammals.
Another type of phylogenetically significant transformations of phylogeny is phylembryogenesis. They represent deviations from ontogenesis characteristic of ancestors, manifested in embryogenesis, but having adaptive significance in adult forms. Thus, hair buds appear in mammals at very early stages of embryonic development, but the hair itself is important only in adult organisms.
Such changes in ontogenesis, being useful, are fixed by natural selection and reproduced in subsequent generations. These changes are based on the same mechanisms that cause congenital malformations: impaired cell proliferation, movement, adhesion, death or differentiation. However, just like cenogenesis, they are distinguished from vices by their adaptive value, i.e. usefulness and fixation by natural selection in phylogenesis.
Depending on the stages of embryogenesis and morphogenesis of specific structures, developmental changes that have the significance of phylembryogenesis occur, three types are distinguished.
1. Anabolism, or extensions, arise after the organ has practically completed its development, and are expressed in the addition of additional stages that change the final result.
Anabolisms include such phenomena as the acquisition of a specific body shape by flounder only after the egg hatches into a fry, indistinguishable from other fish, as well as the appearance of curvatures of the spine, fusion of sutures in the brain skull, and the final redistribution of blood vessels in the body of mammals and humans.
2. Deviations - deviations that arise in the process of organ morphogenesis. An example is the development of the heart in the ontogenesis of mammals, in which it recapitulates the tube stage, two-chamber and three-chamber structure, but the stage of formation of an incomplete septum, characteristic of reptiles, is replaced by the development of a septum, constructed and located differently and characteristic only of mammals. In the development of the lungs in mammals, recapitulation of the early stages of the ancestors is also found, and later morphogenesis proceeds in a new way.
3. Archallaxis - changes that are detected at the level of primordia and are expressed in a violation of their division, early differentiation, or in the appearance of fundamentally new primordia. A classic example of archallaxis is
the development of hair in mammals, the anlage of which occurs at very early stages of development and from the very beginning differs from the anlage of other vertebrate skin appendages.
According to the type of archallaxis, the notochord appears in primitive skullless fish, the cartilaginous spine in cartilaginous fish, and the nephrons of the secondary kidney develop in reptiles.
It is clear that during evolution due to anabolism in the ontogenies of descendants, the basic biogenetic law is fully realized, i.e. recapitulations of all ancestral stages of development occur. In case of deviations, early ancestral stages recapitulate, and later ones are replaced by development in a new direction. Archallaxis completely prevent recapitulation in the development of these structures, changing their very rudiments.
If we compare the diagram of phylembryogenesis with the table of K. Baer, illustrating the law of germinal similarity, it becomes clear that Baer was already very close to the discovery of phylembryogenesis, but the absence of an evolutionary idea in his reasoning did not allow him to be more than 100 years ahead of scientific thought.
In the evolution of ontogenesis, anabolism most often occurs as phylembryogenesis, which only to a small extent changes the entire development process. Deviations as violations of the morphogenetic process in embryogenesis are often rejected by natural selection and are therefore much less common. Archallaxis occur most rarely in evolution due to the fact that they change the entire course of embryogenesis, and if such changes affect the rudiments of vital organs or organs that are important embryonic organizational centers, then they often turn out to be incompatible with life.
In the same phylogenetic group, evolution in different organ systems can occur due to different phylembryogenesis.
Thus, in the ontogenesis of mammals, all stages of the development of the axial skeleton in the vertebrate subtype are traced (anabolism), in the development of the heart only early stages recapitulate (deviation), and in the development of skin appendages there is no recapitulation at all (archallaxis). Knowledge of the types of phylembryogenesis in the evolution of organ systems of chordates is necessary for a doctor to predict the possibility of the occurrence of congenital malformations of an atavistic nature in fetuses and newborns. Indeed, if in an organ system evolving through anabolism and deviation, atavistic developmental defects are possible due to the recapitulation of ancestral states, then in the case of archallaxis this is completely excluded.
In addition to cenogenesis and phylembryogenesis, deviations in the time of organ formation can also be found in the evolution of ontogenesis - heterochrony - and places of their development - heterotopia. Both the first and second lead to changes in the mutual correspondence of developing structures and undergo strict control of natural selection. Only those heterochronies and heterotopies that prove useful are preserved. Examples of such adaptive heterochronies are shifts in the time of the formation of the most vital organs in groups evolving according to the type of arogenesis. Thus, in mammals, and especially in humans, the differentiation of the forebrain significantly advances the development of its other parts.
Heterotopies lead to the formation of new spatial and functional connections between organs, ensuring their further joint evolution. Thus, the heart, located under the pharynx of fish, ensures efficient flow of blood into the gill arteries for gas exchange. Moving to the retrosternal region in terrestrial vertebrates, it develops and functions in a single complex with new respiratory organs - the lungs, performing here primarily the function of delivering blood to the respiratory system for gas exchange.
Heterochronies and heterotopies, depending on at what stages of embryogenesis and organ morphogenesis they appear, can be regarded as phylembryogenesis of different types. Thus, the movement of the rudiments of the brain, leading to its bending, characteristic of amniotes, and manifested at the initial stages of its differentiation, is archallaxis, and heterotopia of the testis in humans from the abdominal cavity through the inguinal canal to the scrotum, observed at the end of embryogenesis after its final formation, - typical anabolia.
Sometimes processes of heterotopy, identical in results, can be phylembryogenesis of different types. For example, in various classes of vertebrates, movement of the limb girdles is very common. In many groups of fish leading a bottom-dwelling lifestyle, the ventral fins (hind limbs) are located anterior to the pectoral fins, and in mammals and humans, the shoulder girdle and forelimbs in the definitive state are located significantly caudal to the place of their initial formation. In this regard, the innervation of the shoulder girdle in them is carried out by nerves connected not with the thoracic, but with the cervical segments of the spinal cord. In the fish mentioned above, the pelvic fins are innervated not by the posterior trunk nerves, but by the anterior segments, located anterior to the centers of innervation of the pectoral fins. This indicates heterotopia in the formation of fins already at the stage of the earliest rudiments, while the movement of the anterior girdle of the limbs in humans occurs at later stages, when their innervation has already been fully accomplished. Obviously, in the first case, heterotopia represents archallaxis, while in the second, anabolia.
Cenogenesis, phylembryogenesis, as well as heterotopia and heterochrony, having proven useful, are fixed in the offspring and reproduced in subsequent generations until new adaptive changes in ontogenesis displace them and replace them. Thanks to this, ontogenesis not only briefly repeats the evolutionary path traversed by our ancestors, but also paves new directions for phylogeny in the future.
Cenogenesis(from the Greek kainós - new and ... genesis (See ... genesis) an adaptation of an organism that occurs at the embryonic (fetal) or larval stage and does not persist in the adult. Examples of c. are the mammalian placenta, which provides the fetus with breathing, nutrition, and excretion; external gills of amphibian larvae; egg tooth in birds, used by chicks to break through the egg shell; organs of attachment in ascidian larvae, swimming tail in trematode larvae - cercariae, etc. The term “C.” introduced in 1866 by E. Haeckel to designate those characteristics that, disrupting the manifestations of palingenesis , i.e., repetitions of distant stages of phylogenesis in the process of embryonic development of an individual, do not allow us to trace the sequence of stages of the phylogeny of their ancestors during the ontogeny of modern forms, i.e., they violate the Biogenetic Law. At the end of the 19th century. Any change in the course of ontogenesis characteristic of the ancestors began to be called color (German scientists E. Mehnert, F. Keibel, and others). Modern understanding of the term “C.” was formed as a result of the work of A. N. Severtsov, who retained for this concept only the meaning of provisional devices, or embryo-adaptations. See also Philembryogenesis.
Cenogenesis(Greek kainos new + genesis origin, formation) - the appearance in the embryo or larva of adaptations to living conditions that are not characteristic of the adult stages, for example. formation of membranes in the embryos of higher animals.
or so
PHYLEMBRYOGENESIS (from the Greek phylon - genus, tribe, embryon - embryo and genesis - origin), an evolutionary change in the ontogenesis of organs, tissues and cells, associated with both progressive development and reduction. The doctrine of phylembryogenesis was developed by the Russian evolutionary biologist A.N. Severtsov. The modes (methods) of phylembryogenesis differ in the time of occurrence during the development of these structures.
If the development of a certain organ in descendants continues after the stage at which it ended in the ancestors, anabolia occurs (from the Greek anabole - rise) - an extension of the final stage of development. An example is the formation of a four-chambered heart in mammals. Amphibians have a three-chambered heart: two atria and one ventricle. In reptiles, a septum develops in the ventricle (first anabolia), but in most of them this septum is incomplete - it only reduces the mixing of arterial and venous blood. In crocodiles and mammals, the development of the septum continues until the complete separation of the right and left ventricles (second anabolia). In children, sometimes, as an atavism, the interventricular septum is underdeveloped, which leads to a serious illness requiring surgical intervention.
Prolonging the development of an organ does not require profound changes in the previous stages of its ontogenesis, therefore anabolism is the most common method of phylembryogenesis. The stages of organ development preceding anabolism remain comparable to the stages of ancestral phylogeny (i.e., they are recapitulations) and can serve for its reconstruction (see Biogenetic Law). If the development of an organ at intermediate stages deviates from the path along which its ontogenesis took place in its ancestors, deviation occurs (from Late Latin deviatio - deviation). For example, in fish and reptiles, scales appear as thickenings of the epidermis and the underlying connective tissue layer of the skin - the corium. Gradually thickening, this anlage bends outward. Then in fish the corium ossifies, the forming bone scales pierce the epidermis and move to the surface of the body. In reptiles, on the contrary, bone is not formed, but the epidermis becomes keratinized, forming the horny scales of lizards and snakes. In crocodiles, the corium can ossify, forming the bony basis of the horny scales. Deviations lead to a more profound restructuring of ontogenesis than anabolism, so they are less common.
Changes in the primary rudiments of organs - archallaxis (from the Greek arche - beginning and allaxis - change) occur least often. In case of deviation, recapitulation can be traced from the origin of the organ to the moment of developmental deviation. In archallaxis there is no recapitulation. An example is the development of vertebral bodies in amphibians. In fossil amphibians - stegocephalians and in modern tailless amphibians, the vertebral bodies are formed around a chord of several, usually three on each side of the body, separate anlage, which then merge to form the vertebral body. In tailed amphibians these anlages do not appear. Ossification grows above and below, covering the notochord, so that a bone tube is immediately formed, which, thickening, becomes the vertebral body. This archallaxis is the reason for the still debated question of the origin of tailed amphibians. Some scientists believe that they descended directly from lobe-finned fish, regardless of other land vertebrates. Others say that tailed amphibians diverged very early from other amphibians. Still others, neglecting the development of the vertebrae, prove the close relationship of tailed and tailless amphibians.
The reduction of organs that have lost their adaptive significance also occurs through phylembryogenesis, mainly through negative anabolism - loss of the final stages of development. In this case, the organ either underdevelops and becomes a rudiment, or undergoes reverse development and completely disappears. An example of a rudiment is the human appendix - an underdeveloped cecum; an example of complete disappearance is the tail of frog tadpoles. Throughout its life in water, the tail grows, new vertebrae and muscle segments are added at its end. During metamorphosis, when the tadpole turns into a frog, the tail dissolves, and the process occurs in the reverse order - from the end to the base. Phylembryogenesis is the main method of adaptive changes in the structure of organisms during phylogenesis.
Ticket 97
Principles (methods) of phylogenetic transformations of organs and functions. Correspondence between structure and function in living systems. Multifunctionality Quantitative and qualitative changes in the functions of biological structures.
.
Principles of phylogenetic transformations
Authority call a historically developed specialized system of tissues, characterized by delimitation, constancy of shape, localization, internal structure of circulatory and innervation pathways, development in ontogenesis and specific functions. The structure of organs is often very complex. Most of them are multifunctional, i.e. performs several functions simultaneously. At the same time, various organs may participate in the implementation of any complex function. A group of organs similar in origin that unite to perform a complex function is called system(circulatory, excretory, etc.). If the same function is performed by a group of organs of different origin, it is called apparatus. An example is the respiratory apparatus, which consists of both the respiratory organs themselves and the elements of the skeleton and muscular system that provide respiratory movements.
In the process of ontogenesis, development and often replacement of some organs by others occurs. The organs of a mature organism are called definitive; organs that develop and function only in embryonic or larval development - provisional. Examples of provisional organs are the gills of amphibian larvae, the primary kidney, and the embryonic membranes of higher vertebrates (amniotes).
In historical development, organ transformations may have progressive or regressive character. In the first case, the organs increase in size and become more complex in their structure; in the second, they decrease in size and their structure becomes simpler.
If two organisms located at different levels of organization have organs that are built according to a single plan, located in the same place and develop in a similar way from the same embryonic rudiments, then this indicates the relationship of these organisms. Such organs are called homologous. Homologous organs often perform the same function (for example, the heart of fish, amphibians, reptiles and mammals), but in the process of evolution the functions can change (for example, the forelimbs of fish and amphibians, reptiles and birds).
When unrelated organisms live in the same environments, they may develop similar adaptations, which manifest themselves in the appearance similar organs. Similar organs perform the same functions, but their structure, location and development are sharply different. Examples of such organs are the wings of insects and birds, the limbs and jaw apparatus of arthropods and vertebrates.
The structure of organs strictly corresponds to the functions they perform. Moreover, in the historical transformations of organs, a change in functions is certainly accompanied by a change in the morphological characteristics of the organ.
The basic principle of the evolution of organic structures is the principle differentiation. Differentiation is the division of a homogeneous structure into separate parts, which, due to different positions, connections with other organs and various functions, acquire a specific structure. Thus, the complication of the structure is always associated with the complication of functions and specialization of individual parts. The differentiated structure performs several functions, and its structure is complex.
An example of phylogenetic differentiation can be the evolution of the circulatory system in the chordate phylum. Thus, in representatives of the skullless subtype it is structured very simply: one circle of blood circulation, the absence of a heart and capillaries in the system of gill arteries.
The superclass of fish has a two-chambered heart and gill capillaries. In amphibians, for the first time, the circulatory system is divided into two circulation circles, and the heart becomes three-chambered. Maximum differentiation is characteristic of the circulatory system of mammals, whose heart is four-chambered, and in the vessels complete separation of the venous and arterial blood flows is achieved.
Individual parts of a differentiating, previously homogeneous structure, specializing in performing one function, become functionally more and more dependent on other parts of this structure and on the organism as a whole. This functional subordination of individual components of the system in the whole organism is called integration.
The four-chambered heart of mammals is an example of a highly integrated structure: each department performs only its own special function, which has no meaning in isolation from the functions of other departments. Therefore, the heart is equipped with an autonomous system of functional regulation in the form of the parasympathetic atrioventricular nerve node and at the same time is strictly subordinate to the neurohumoral regulation system of the body as a whole.
Thus, simultaneously with differentiation, the subordination of parts to the integral system of the body is also observed, i.e. integration process.
In the process of evolution, it is natural that emergence new structures and their disappearance. It is based on the principle of differentiation, which manifests itself against the background of primary polyfunctionality and the ability of functions to change quantitatively. Any structure in this case arises on the basis of previous structures, regardless of at what level of organization of living things the process of phylogenesis takes place. Thus, it is known that about 1 billion years ago, the original globin protein, following duplication of the original gene, differentiated into myo- and hemoglobin - proteins that are part of muscle and blood cells, respectively, and therefore differentiated by function. In the phylogenesis of the central nervous system of chordates, one can also see the differentiation and change in functions of structures: the brain is formed from the anterior end of the neural tube. In the same way, new biological species are formed in the form of isolated populations of the original species (see § 11.6), and new biogeocenoses - due to the differentiation of pre-existing ones (see § 16.2).
Due to the fact that the phylogenies of specific organ systems will be considered below, we will dwell in more detail on the patterns of the appearance and disappearance of organs. Example emergence organs is the origin of the uterus of placental mammals from paired oviducts. With the lengthening of the embryonic development of mammals, the need arises for a longer retention of the embryo in the mother's body. This can only be carried out in the caudal sections of the oviducts, the cavity of which is enlarged, and the wall is differentiated in such a way that the placenta is attached to it, ensuring the relationship between the mother and the fetus. In the process of natural selection, those mammals in whose bodies the offspring developed for the longest time were preserved and successfully reproduced first of all. As a result, a new organ emerged - the uterus, which provides the embryo with optimal conditions for intrauterine development (see Section 14.5.3) and increases the survival rate of the corresponding species.
In the emergence of such a more complex and specialized organ as the eye, the same patterns are observed. The formation of the organ of vision, like all sense organs, is based on skin epithelial cells, among which receptor cells, in particular photosensitive cells, are differentiated. Combining them into groups leads to the emergence of primitive separate organs of vision, allowing animals only to evaluate illumination. Immersion of such a light-sensitive organ under the skin ensures the safety of delicate cells, but visual function can only be achieved due to the appearance of transparency of the integument. The sensitivity to light of the primitive organ of vision increases with the thickening of the transparent covers and their acquisition of the ability to refract light and focus its rays on the sensitive cells of the eye. A complex organ requires an auxiliary apparatus - protective structures, muscles that move it, etc. The increased level of complexity of the organization of the eye is necessarily accompanied by a complication of the regulation of its functions, which is expressed in the strengthening of its integration as an integral system.
Disappearance or reduction of an organ in phylogeny can be associated with three different reasons and has different mechanisms. Firstly, an organ that previously performed important functions may turn out to be harmful in new conditions. Natural selection works against it, and the organ can completely disappear quite quickly. There are few examples of such direct disappearance of organs. Thus, many insects of small oceanic islands are wingless due to the constant elimination of flying individuals from their populations by the wind. The disappearance of organs is more often observed due to their substitution by new structures that perform the same functions with greater intensity. Thus, for example, in reptiles and mammals, the forebuds and primary kidneys disappear, being replaced functionally by secondary kidneys. In the same way, in fish and amphibians, the notochord is displaced by the spine.
The most common path to the disappearance of organs is through the gradual weakening of their functions. Such situations usually arise when living conditions change. An organ that performs almost no functions escapes the control of natural selection and usually exhibits increased variability. The resulting changes cause disruption of correlative connections with other parts of the body. Due to this, such an organ often becomes harmful and natural selection begins to act against it.
It is widely known in medical practice that rudimentary organs in humans are characterized by wide variability. Third large molars, or “wisdom teeth,” for example, are characterized not only by significant variability in structure and size, but also by different periods of eruption, as well as a special susceptibility to caries. Sometimes they do not erupt at all, and often, having erupted, they are completely destroyed over the next few years. The same applies to the vermiform appendix of the cecum (appendix), which normally can have a length from 2 to 20 cm and be located differently (behind the peritoneum, on the long mesentery, behind the cecum, etc.). In addition, inflammation of the appendix (appendicitis) occurs much more often than inflammatory processes in other parts of the intestine.
The process of organ reduction is opposite to its normal morphogenesis. First of all, the bookmarks of such parts of the organ fall out, which are normally the last to form. When a person’s limbs are underdeveloped, the phalanges of the 1st and 5th fingers, which are the last to develop, are usually the first to be underdeveloped. In cetaceans, which are completely devoid of hind limbs due to the weakening of their functions in phylogenesis, the rudiments of the elements of the pelvic girdle still remain, which are formed very early in the process of morphogenesis.
Studies of the genetic basis of organ reduction have shown that structural genes regulating morphogenesis do not disappear, while genes regulating the time of formation of rudimentary organs, or genes responsible for the phenomenon of inductive interactions in the developing embryo, undergo significant changes. Indeed, when the mesodermal material of the floor of the oral cavity of a lizard embryo is transplanted into the oral cavity of a developing chick, it is possible for the latter to form teeth of a typical structure, and the transplantation of the skin mesoderm of a lizard under the epidermis of the back of a chick leads to the formation of typical horny scales in it instead of feathers.
Similarities between embryos of representatives of different groups of vertebrates
The development of all animals begins with one cell.
Organogenesis occurs in a certain sequence. In chordates it begins With formation of the rudiment of the notochord and nervous system. On the dorsal side of the embryo, a group of ectoderm cells separates into a long plate. These cells begin to actively divide, plunging into the body of the embryo and forming a groove, the edges of which gradually come closer together and then close, forming the primary neural tube.
In addition to the nervous system, skin glands, tooth enamel, hair, nails, and skin epithelium also arise from the ectoderm. . Endoderm gives rise to the tissues lining the intestines and respiratory tract , forms the liver and pancreas. Muscles, cartilage and bone skeleton, excretory organs are formed from the mesoderm , reproductive and circulatory systems of the body .
During the process of embryogenesis, there is a close interaction between the parts of the developing embryo: the rudiment of one organ or organ system determines (induces) the location and time of formation of another organ or organ system.
Differentiation of embryonic cells does not occur immediately, but at a certain stage of development. In the early stages of cleavage, the cells of the embryo are not yet specialized, so each of them can give rise to a whole organism. If for any reason these cells separate, two identical cells are formed. embryo, containing identical genetic information, each of which develops into a full-fledged individual. As a result, identical or monozygotic twins are born. In the human population, these are the only people who have an identical genotype and are copies of each other,
The mutual influence of the parts of the embryo has been demonstrated in numerous experiments. German researchers Hans Spemann and Hild Mangold took from newt embryos at the early gastrula stage a section of the dorsal side of the body, from which V further, the notochord and mesoderm should have developed, and it was transplanted to the ventral side of another gastrula . IN As a result, on the abdominal side of the second embryo, an additional neural tube was formed from the cells that were supposed to give rise to the skin. This phenomenon is called Embryonic induction.
Creation of comparative embryology
K.M. Bare was the first to apply the embryological criterion in systematics and substantiate its significance. Comparing the methods of animal development, the scientist came to the conclusion that there are 4 types of organization - the vertebrate type, the elongated or articulated type, the massive type and the peripheral or radiant type. Each type corresponds to a special development plan. Within each type, different levels of development can be distinguished, corresponding to classes. Animals differ from each other both in the type of ontogenesis and in the degree of heterogeneity.
Thus, by the middle of the 19th century, on the one hand, differences in the ontogenesis of animals of different types and classes were outlined. On the other hand, it became clear that the embryonic development of very dissimilar organisms proceeds with the formation of germ layers, which allows us to talk about homologous structures in representatives of different groups. It became possible to compare data from embryology and comparative anatomy.
At the beginning of the 19th century. I. Meckel, L. Oken, M. Rathke, Gidemman put forward the assertion that the embryos of higher animals go through in their development all stages corresponding to the adult state of lower animals.
K.M. Bare, having carefully studied the individual development of representatives of different groups of vertebrates and invertebrates, came to the conclusion that only animal embryos can be compared. In the process of embryonic development, the characters of the phylum appear first, then the class, order, family, genus and species. The younger the embryos belonging to different classes of the same type, the more similar they are to each other. In other words, representatives of different groups, for example, classes of the vertebrate subtype, in the early stages of ontogenesis are usually more similar than adult individuals. This pattern is called Law of germinal similarity .
The results of embryological studies by K.M. Baer were used C. Darwin as one of the proofs of evolution.
Comparative embryology
Von Baer (1792-1867), studying embryonic development in representatives of different groups of vertebrates, discovered surprising structural similarities in all these groups, especially at the stages of cleavage, gastrulation and differentiation of the nascent organism.
Haeckel (1834-1919) suggested that this similarity had evolutionary significance. He formulated the law of recapitulation, according to which “ Ontogenesis repeats phylogeny", i.e., the stages through which an organism passes in the process of its development repeat the evolutionary history of the group to which it belongs. The study of the early embryos of any vertebrates alone shows that it is impossible to determine the group to which they belong. It is only at relatively late stages of development that the embryo begins to bear some resemblance to the corresponding adult form. The initial similarity between embryos is explained by the fact that they all, and therefore the classes to which they belong, had a common ancestor. The law of recapitulation, however, cannot be accepted unconditionally, since not a single living organism can exhibit all the signs of its putative evolutionary ancestors. But it seems likely that organisms retain developmental mechanisms inherited from their ancestors. Therefore, it is possible that at different stages of development a given organism will have features of structural similarity with the embryos of ancestral forms. Subsequent adaptations to other environmental conditions and lifestyles change the further course of development. As observations show, the closer the groups to which two given organisms are assigned on the basis of common homological structures, the longer their similarity remains at the embryonic stages. Organisms adapted to a certain way of life and a certain habitat, not typical for the large group to which they belong, are less similar to other members of this group in the process of embryonic development.
BER'S LAW- a group of provisions postulating the similarity of the early stages of development of embryos of representatives of different classes of vertebrate animals. K. E. Baer (a Russian embryologist who lived in the 19th century) believed that first, embryos develop characters of a type, then a class, an order, and, finally, in the later stages of development - individual characters that characterize the specialization of the taxon, i.e. embryos of different classes are at first similar, and then deviate from each other in their development.
MUELLER-HOCKEL LAW (biogenetic) - states that ontogeny is a brief repetition of phylogeny. The law was formulated by E. Haeckel based on the ideas of Charles Darwin and the research of F. Muller. At the beginning of the 20th century. the law was supplemented by the position of A. N. Severtsov that in the ontogenesis of organisms, not entire phylogenetic stages are repeated, but only recapitulation (short and condensed repetition) of individual characteristics and processes is possible.
Law of germinal similarity
Explorers of the early 19th century. for the first time they began to pay attention to the similarity of the stages of development of embryos of higher animals with the stages of complexity of organization leading from lowly organized forms to progressive ones. Comparing the stages of development of embryos of different species and classes of chordates, K. Baer made the following conclusions.
1. Embryos of animals of the same type are similar in the early stages of development.
2. They consistently move in their development from more general characteristics of the type to more and more specific ones. Lastly, signs develop that indicate that the embryo belongs to a certain genus, species, and, finally, individual traits.
3. Embryos of different representatives of the same type gradually become separated from each other
Similarities of embryos and embryonic divergence of characters (K. Baer’s law)
biogenetic law (E. Haeckel and F. Müller) by A. N. Severtsev
1. Introduction
4. Works of Severtsov A.N.
Bibliography
1. Introduction
Natural philosophers of the 18th and 19th centuries. They said that life is fundamentally one. This unity was expressed in their parallelism between the embryonic development of an individual and the ladder of living beings, which dates back to Aristotle. According to the concept of the ladder of living beings, all objects existing in nature are links in a continuous chain connecting inorganic creations with a series of living forms of increasing complexity. From inanimate nature there is a gradual transition to plants, and then to such simple animals as sponges, then to insects, fish, birds, mammals, and finally to humans. This pattern was static and should be interpreted as evolutionary. It simply represented the plan according to which God created the world.
According to the law of parallelism, known as the Meckel-Serres law (after the names of its two creators: Meckel in Germany, Serres in France), every living creature in its embryonic development repeats the adult forms of animals standing on lower steps of the ladder of living beings. Conversely, lower animals represent permanent personality stages of evolutionarily more advanced forms. The law of parallelism, like the ladder of living beings, did not contain anything evolutionary. A detailed analysis of such parallelism allowed Karl Baer to formulate the law of germinal similarity.
2. Similarities of embryos and embryonic divergence of characters (K. Baer’s law)
Karl Maksimovich Baer (aka Karl Ernst von Baer) was born in 1792 in Estland, and here, in Dorpat (now Tartu), he completed his life’s journey in 1876. The Konigsberg period of Baer's activity is associated with embryology. Working at the University of Königsberg, he conducted his research, which largely laid the foundations of embryology as a science, in an atmosphere of dominance of natural philosophy.
He, following Wolf and Pander, studied the embryonic development of the chicken, and then a number of other representatives of different classes of vertebrates. He discovered the egg in mammals, studied the development of fetal membranes and germ layers; he discovered and described the blastula, a stage of embryonic development in animals. All these discoveries were made by him at the age of 25-36 years. Baer spent days and nights, weeks and months in the laboratory.
What fascinated the young researcher so much? Why did he, in his words, “turn into some kind of hermit crab, never leaving the shell he had once chosen”? Baer was able to take a fresh look at the long-known phenomenon of similarity in the embryonic stages of animals belonging to different classes of vertebrates.
Baer was the first to identify three forms of differentiation in embryology: splitting into germ layers, or “primary separation”; the emergence of tissues, or “pure-logical isolation” (this was written a decade and a half before the creation of the cell theory!); the emergence of organs, or “morphological separation”. Based on the results of his work on comparative embryology, he made a number of generalizations that showed the complete meaninglessness of the idea that animals in their development repeat all the steps of the ladder of living beings.
Baer, like Cuvier, who in 1812 created a classification system that crossed out the ladder of living beings, noticed that there is not one sequential series, but four basic plans for the structure of animals. These four planes are clearly reflected in their development. For example, the notochord* and neural tube characteristic of vertebrates arise in the early stages of development and thus “the embryo of a vertebrate animal is from the very beginning a vertebrate animal, and no period corresponds to an invertebrate animal.” Vertebrate embryos are similar only to other vertebrate embryos; Baer denies their resemblance to the adults of any other animals: “The embryos of Vertebrata* do not pass through the permanent forms of any (known) animals in the process of their development.” Facts, not preconceived notions, led him to evolutionary views.
Pay attention to how modern the scientist’s idea sounds that “the more different life forms are from each other, the further into the depths of the history of development one must dive in order to find similarities between them.” However, by the words “history of development” Baer meant the history of individual, that is, embryonic, and not historical development.
Summarizing the comparative data, Baer published in 1828 the following basic generalizations - his famous laws, diametrically opposed to the theory of parallelism:
1. The common of each larger group of animals is formed in the embryo earlier than the special...
2. From the more general... the less general is formed, and so on, until, finally, the most special arises...
3. The embryo of each animal form does not at all repeat other animal forms during development, but, on the contrary, rather separates itself from them.
4. Basically, then, the embryo of the highest form of an animal is never similar to another form of animal, but only to its embryo.
It is not difficult to notice the polemical overtones of Baer's conclusions, especially the last two, directed against the theory of parallelism. However, they were supported by facts, and two main provisions, called the principle of embryonic similarity and the principle of specialization of individual development, have retained their significance to the present day. Indeed, the embryos of different species are very similar to each other, and as they develop, this similarity is lost. In the early stages of development of the chick embryo - Baer's favorite subject of research - one can only see that it belongs to the vertebrates, because the early embryos of vertebrates of all classes look almost the same; a little later it can be identified as a bird, and only later does it become obvious that this is a future chicken.
The paths and patterns of individual development, the laws of ontogenesis, questions of the relationship between ontogeny and phylogenesis were then developed by Darwin, Müller and Haeckel, A. N. Severtsov. But all of them in their constructions proceeded from the factual foundation that was created in embryology by the works of Karl Baer.
This outstanding scientist is a major figure in natural science of the 19th century. Embryology, ichthyology, theriology, comparative anatomy, geography, ethnography, hunting and fishing, history of science, bibliography - this is not a complete list of those areas of knowledge in each of which Baer left his traces. Baer was not an evolutionist, but with his law he prepared the ground for an evolutionary interpretation of the processes of ontogenesis. Already Charles Darwin emphasized that the study of the individual development of organisms sheds light on their origin, since “in the embryo one can see a vague portrait of an ancestor.” This position formed the basis of the famous biogenetic law established by Ernst Haeckel and Fritz Müller.
3. Biogenetic law (E. Haeckel and F. Muller)
With the advent of Charles Darwin's theory on the origin of species (1859), J. Meckel's theory of parallelism received an evolutionary explanation: the ladder of creatures ceased to be static, it became clear that it reflects the evolutionary tendency to improve organization, and ontogenesis - the individual development of an individual - is a process of evolution. At the same time, it became possible to explain embryonic similarity by kinship (common origin) of organisms, but the principle of specialization of individual development remained unexplained, and K. Baer was an opponent of both I. Meckel and Charles Darwin until the end of his life. Thus, general ideas about ontogenesis turned out to be contradictory: on the one hand, this process reflects the evolution of the organization, on the other, only itself.
In addition, Charles Darwin, to explain the evolution of the early stages of development, developed the principle of inheritance at the appropriate age, according to which a trait, once arising at one stage of ontogenesis, tends to appear in descendants at the same stage, which makes it possible for selection to change any stage of individual development . This pattern did not explain either parallelism or specialization of development. As a result, the need arose to reconcile all the contradictory concepts, and the study of the relationship between individual and historical development was started almost anew.
Darwin's ideas were developed by Fritz Müller, who did a lot in substantiating the biogenetic law. F. Müller (1821-1897) - German zoologist, who left for Brazil forever in 1852. He never met either Darwin or Haeckel. From the provincial Brazilian town of Desterro, Müller sent a small book entitled “For Darwin” (1864) for publication in Leipzig. In it, based on studies of the ontogeny of higher crustaceans, he came to two important conclusions:
1. The ontogeny of descendants can continue beyond the stage at which it ended in the ancestors. Then the organization of the descendants becomes higher than that of their ancestors.
2. The ontogeny of descendants may, at intermediate stages, deviate away from the path along which it followed among their ancestors. Then the organization of the descendants will be different, but not higher than that of their ancestors.
In this work, neither the mechanisms of individual development nor the evolution of ontogenesis as a process were discussed. We were talking only about the evolution of the adult stage, and we considered how changes in the course of ontogenesis affect the definitive structure. The reasons for changes in the course of ontogenesis were not touched upon.
And yet this work of F. Muller was an important step forward in the development of the theory of the evolution of ontogenesis. Firstly, he showed that evolutionarily new forms arise by changing the course of individual development characteristic of their ancestors, i.e. changes in ontogeny are primary in relation to phylogenetic changes. Secondly, it was shown that changes in the course of ontogenesis can occur in different ways. Thirdly, based on Müller’s data, it can be argued that if the ontogenesis of descendants continues beyond the definitive stage of the ancestors, from this, which has already become the penultimate, stage, one can judge the organization of the ancestors and thereby the continuation of the studied forms. As a result, the theory of parallelism was partially explained and the role of embryology in the reconstruction of phylogeny was substantiated. It should only be noted that Muller considered the ontogeny of the whole organism, and not its parts (individual organisms and histogenesis*).
F. Muller's ideas served as the basis for further development of the problem. They were used by Ernst Haeckel (1834-1919), who played a huge role in the propaganda and further development of evolutionary teaching. Haeckel and Müller developed the idea that successive changes in the shape of an individual during individual development are due to phylogenesis, that is, the development of the genus to which a given species of animal belongs.
Ontogenesis is a short and rapid repetition of phylogeny, due to heredity and adaptation. This pattern was called the basic biogenetic law by Haeckel (1866). A more complete formulation of this law is as follows: the development of the embryo (individual or ontogenetic series of development, ontogenesis) is a compressed, abbreviated repetition of the development of the entire group of forms that arose from one common root (i.e., a repetition of the development of phyletic, or paleontological, phylogeny); the more complete the repetition is, the more the original development of the ancestors (palingenesis) is preserved due to constant inheritance. The repetition is less complete, the more later disturbances (coenogenesis) were introduced thanks to various adaptations.
According to this law, the embryos of higher animals are similar to the adult lower ancestors. The eggs of all animals are "identical". Evolution occurs through the addition of new stages at the end of development. If nothing interfered, then the evolutionary history of the species would exactly repeat in individual development, which means that during the hours, weeks or months of development of the embryo of this or that animal, we could watch, figuratively speaking, a “movie” about its historical past. When watching a “movie” about the individual development of mammals, we would see stages corresponding to the adult forms of invertebrate animals, and then fish, amphibians, and reptiles.
However, the manifestation in an unchanged form of primitive, ancient, palingenetic characters (from the Greek palaios - ancient; palingenesis - characters that passed into the ontogenesis of an animal from its phylogeny) is hampered by cenogenesis, various characters that arose in ontogenesis as an adaptation to the living conditions of embryos of larvae and adult animals . Examples of coenogenesis include the amnion, chorion, allantois of Amniota, and the huge spinning glands of insect larvae.
The process of repetition of phylogenetic characters in ontogenetic development is called recapitulation. A typical example of recapitulation is given by Academician A. N. Severtsov in his book “Morphological Patterns of Evolution,” published in 1913 in Germany. We know that in modern tailless amphibians, the tibia and fibula are fused together as adults, but in tadpoles they are separated. Stegocephalians, from which modern amphibians evolved, also had two separate shin bones. Consequently, the presence of separate tibia bones in tadpoles can be considered as a recapitulation of one of the characteristics of the hind limbs characteristic of the ancestral skeleton.
Recapitulation is not limited to morphological characteristics. They can also be identified during the ontogenetic development of the functions of various organs and tissues. It is known that during the evolution of vertebrates, the enzymes necessary for the breakdown of uric acid were gradually lost. Thus, in some reptiles and birds the final product of nitrogen metabolism is uric acid, in amphibians and most fish it is urea, and in primary invertebrates it is ammonia. It turned out that the bird embryo secretes ammonia in the early stages of development, urea in the later stages, and uric acid only in the last stages. Similarly, in tadpoles the end product of metabolism is ammonia, and in frogs it is urea.
The biogenetic law became the core of Haeckel's theory of recapitulation. According to this theory, ontogenesis includes two types of characters: palingenesis - characters of phylogenetic distant ancestors and coenogenesis - secondary characters that arose as a result of adaptation to certain conditions of embryonic and larval life. Typical coenogenesis are, for example, the tracheal gills of dragonfly and mayfly larvae: in an aquatic lifestyle, they serve as temporary respiratory organs.
Haeckel attached a primary role to palingenetic characters inherited from distant ancestors in elucidating phylogenetic relationships. The study of cenogenesis in this aspect is useless and only obscures the manifestation of palingenetic characteristics. From this point of view, cenogeneses are “annoying distortions” of palingenesis.
There were many omissions in Haeckel's theory, and the main one was that the history of the adult organism was considered in isolation from the history of the embryo. Despite this, one cannot underestimate the enormous importance of the biogenetic law in the development of evolutionary teaching. With further research, however, it turned out to be increasingly difficult to fit the accumulating facts into the Procrustean bed of a narrowly formulated theory. Criticism intensified, sometimes reaching the point of completely denying the biogenetic law and the very principle of recapitulation. It seemed that he had exhausted himself and was moving into the field of the history of science with its inevitable mistakes.
4. Works of A. N. Severtsov
Academician Alexei Nikolaevich Severtsov (1866-1936) managed to overcome these incorrect views. The theory of phylembryogenesis he proposed restored the biogenetic law and the principle of recapitulation, giving them a completely new light. A. N. Severtsov developed his theory all his life.
It should be noted that A. N. Severtsov and his school concentrated their attention on the laws of macroevolution. Severtsov's earliest general biological speech - the speech “Evolution and Embryology” - dates back to 1910; in 1922 he published “Studies on the Theory of Evolution. Individual development and evolution". These works provide criticism of the biogenetic law in its Haeckelian mechanistic form, based on specific research by A. N. Severtsov and his school. Discussing the eternal problem of adaptability and neutrality of characters, Severtsov proposed to distinguish between primary adaptive changes (prothallaxis) and secondary changes that occurred due to correlative variability (deutalaxis). Severtsov drew attention to the existence of phylogenetic changes associated with changes in the course of embryonic development - he called such adaptations phylembryogenesis.
Speaking about the relationship between onto- and phylogeny, Severtsov wrote: “The biogenetic law of Müller-Haeckel cannot be considered refuted - it contains a very significant amount of truth, although not the complete truth: the relationship between phylogeny and ontogenesis is much more complex than hitherto thought , and are not limited to the relations discovered by Müller and developed by Haeckel.” Severtsov pointed out the differences in the timing of the appearance of new characters in ontogenesis and established that new characters do not always “supercharge” a number of successive stages that occur in ontogenesis. In some cases this series is preserved, while in others the entire sequence of stages of ontogenesis may be transformed.
“Morphological patterns of evolution” is the main theoretical work of A. N. Severtsov. It was published in Russia in 1939, after the death of the scientist. Possessing vast experience in comparative anatomical and embryological research, Severtsov gave in this book a broad picture of macroevolution, its patterns and methods of study.
In the first chapters, he examined the tasks and methods of phylogenetic research and described the theory of monophyletic (single-root) evolution as satisfying factual and theoretical constructs. Despite the obvious bias towards the study of the evolution of higher taxa, Severtsov also paid attention to the adaptability of characters of small systematic groups. He considered the existence of selectively indifferent characters possible, but believed that they spread in the population due to their correlation with selectively essential characters. 6 years before the publication of Theodosius Grigorievich Dobzhansky’s book “Genetics and the Origin of Species,” A. N. Severtsov gave a close definition of species: we unite similar animals into one species based on a biological trait, that is, based on their ability to mate with each other and produce fertile offspring "
Changes in the individual development of organisms or in the development of certain organs, persistently transmitted to offspring, new characteristic features can arise at the beginning of embryonic development, at the beginning of the formation of the corresponding organs - Severtsov came to this conclusion. This type of phylembryogenesis is called archallaxis. In other cases, Severtsov noted, changes may affect any middle stages in the development of organs (or the entire organism). This type of change is called deviation. Finally, changes could occur at the final stages of the development of organisms or at the final stages of organ development. These changes are called anabolism (i.e., a transition to the final stages of development).
Thus, according to Severtsov, various characteristics of an adult animal are formulated at different times during its ontogenesis and the ontogenesis of organs, and then pass unchanged into the adult state as definitive characteristics; additions occur over the definitive state of characters, which, developing in ontogenesis, is both “embryonic” and “adult” at the same time.
Severtsov created a hypothesis of the origin and evolution of the ontogeny of multicellular animals, and the ideas about anabolism and recapitulations based on them are of great importance in it. Being a supporter of hypotheses about the origin of multicellular organisms from unicellular colonies, Severtsov suggests that in the ancestors of modern multicellular organisms, namely those that are similar to the living plants Pandorina morum or Volvox, ontogenesis was short and consisted of superimposed stages during the development of colonies; from the one-cell stage a two-cell (first superstructure) evolved, from a two-cell stage a four-cell stage (second superstructure), etc.
In Volvox, through a series of such supers, the organism develops to a spherical colony, which can consist of more than 20,000 cells, then a stage (super) of pure differentiation occurs (of a certain variety of cell types). In animals such as lower coelenterates, ontogenesis has lengthened. Hydra, according to Severtsov, has stages in ontogenesis corresponding to the stages of ontogenesis of colonial forms - a period of fragmentation. To these stages are added the stages of the formation of a two-layer embryo - gastrula, then the formation of a polyp with an ectoderm, gastric cavity, and tentacles. To such “superstructure” stages is added only the stage of growth and pure differentiation (formation of stinging and other cells). Thus, the ontogeny of the hydroid was extended by several new stages added to the definitive state of the colonial forms of unicellular organisms.
Evolutionary changes in ontogenesis that lead to the appearance of new characters are positive phylembryogenesis. There are also negative phylembryogenesis, which results in complete or partial reduction of an organ or its parts. A. N. Severtsov distinguished two types of reductions: rudimentation and aphanesia.
With rudimentation, either a simplification of the organ occurs from the very initial stages of its embryonic development, or, as a result of retardation, a later formation of the organ occurs and, as a result, its underdevelopment. In the latter case, the final stages disappear, and in adult forms this organ takes on a juvenile appearance. An example of such rudimentation is the modified limbs of serpentine lizards: in Seps chalcides only three fingers are formed and the limbs are embryonic in nature.
With the second type of reduction - aphanisia - the organ developed in the ancestors completely disappears in their descendants, but the embryonic anlage is nevertheless preserved. An example of aphanisia is the atrophy of the tail in tadpoles, which disappears during metamorphosis as a result of resorption, in contrast to the ancestors of tailless amphibians, in which the tail was also developed in adult forms. In the same way, the superbly developed teeth in the fry of modern sturgeons completely disappear in adults due to a change in diet; however, in the ancestors of Acipenseridae, teeth were also developed in adult fish.
So, according to the theory of A. N. Severtsov, various changes in ontogenesis lead not only to the appearance of new characters in adult forms, but also to their disappearance on the basis of positive or negative phylembryogenesis.
The variety of adaptive changes in organs, which is formed in a variety of ways, was carefully analyzed by A. N. Severtsov. In his classification of modes (methods) of phylogenetic changes in organs, he proceeded from the functional principle, considering morphological changes in organs as “a way of developing an active function or passive adaptation in a direction useful for the whole organism.” Thus, when speaking about changes in any part of the body, Severtsov took into account the integrity of the entire organism and its connection with environmental conditions.
The theory of phylembryogenesis quite satisfactorily explains the evolution of the structure of organisms at all levels - from cell to individual and at all stages of ontogenesis - from zygote* to adult animal. Any ontogenesis and any of its stages can be considered as a complex system, an interweaving of phylembryogenesis of organs, tissues, and cells. The evolution of individual development occurs through the evolution of particular organo-, pure-, and cytogeneses.
A. N. Severtsov had a large school; his influence was felt not only in our country. Not only F. G. Dobzhansky and N. V. Timofeev-Resovsky, but also B. Rensch and A. Remane were familiar with his ideas. In his later works, A. N. Severtsov turned to genetic data and supported the struggle of geneticists against neo-Lamarckism. However, Severtsov’s works were ignored by such active creators of the synthetic theory of evolution as J. Huxley and J. G. Simpson. The indirect influence of Severtsov’s ideas on Western evolutionists is beyond doubt. Among his many students there was one who went much further in biological science than his teacher. This is I.I. Shmalgauzen, who throughout his life, like a teacher, was engaged in comparative anatomy, embryology and general phylogenetics. He became the successor to the work of A.N. Severtsov and one of the first researchers to carry out the synthesis of classical Darwinism, with all its components, with genetics.
Bibliography
1. Large encyclopedic dictionary. Great Russian Encyclopedia, St. Petersburg, Norint, 2001.
2. Vorontsov N. N. Development of evolutionary ideas in biology. M.: Publishing house. Department of UC DO MSU, Progress-Tradition, ABF, 1999.
3. Reff R., Kofman T. Embryos, genes, evolution, M.: Mir, 1986.
4. Tokin B. P. General embryology. M.: Higher School, 1987.
5. Toshchenko V.P. Introduction to the theory of evolution: a course of lectures /pod. Ed. Polyansko-
6. Oxford Illustrated Encyclopedia, Vol. 2. Natural World. M., Publishing House "Infra-M", "The Whole World", 1999.
7. A. S. Severtsov. Basic theories of evolution. M., Moscow State University Publishing House, 1987
2.1 K. Baer’s law of germline similarity
Explorers of the early 19th century. for the first time they began to pay attention to the similarity of the stages of development of embryos of higher animals with the stages of complexity of organization leading from lowly organized forms to progressive ones. In 1828, Karl von Baer formulated a pattern called Baer's Law: “The earlier stages of individual development are compared, the more similarities can be found.” Comparing the stages of development of embryos of different species and classes of chordates, K. Baer made the following conclusions.
1. Embryos of animals of the same type are similar in the early stages of development.
2. They consistently move in their development from more general characteristics of the type to more and more specific ones. Lastly, signs develop that indicate that the embryo belongs to a certain genus, species, and, finally, individual traits.
3. Embryos of different representatives of the same type gradually become separated from each other (Fig. 1).
Figure 1. Similarity of embryos of different classes of vertebrates at different stages
K. Baer, not being an evolutionist, could not connect the patterns of individual development he discovered with the process of phylogenesis. Therefore, the generalizations he made had the meaning of no more than empirical rules.
The subsequent development of the evolutionary idea made it possible to explain the similarity of early embryos by their historical kinship, and their acquisition of more and more particular traits with gradual isolation from each other - the actual isolation of the corresponding classes, orders, families, genera and species in the process of evolution.
The theory of evolution developed by Charles Darwin clearly highlighted the fundamental importance of the problem of ontogenetic development. Embryonic similarity is now explained by the actual relationship of organisms, and their gradual divergence (embryonic divergence) serves as an obvious reflection of the historical divergence of these forms (phylogenetic divergence). In the embryo of our descendants, Darwin wrote, we see a “vague portrait” of our ancestors. Consequently, the history of a given species can be traced by individual development.
Animal Biology
The similarity between arthropods and annelids lies in such features as body segmentation, abdominal nerve cord, and the presence of a circulatory system. Also related and descended from ancient annelids. Their differences are...
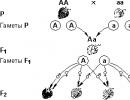
Genetics and evolutionary teaching
Genetics and evolutionary teaching
This law states that crossing individuals that differ in a given trait (homozygous for different alleles) produces genetically homogeneous offspring (F 1 generation), all individuals of which are heterozygous...
Genetics and evolution. Mendel's laws of genetics
This law is called the law of (independent) splitting. Its essence is as follows. When an organism heterozygous for the trait under study develops sex cells - gametes, then one half of them carries one allele of a given gene...
Genetics and evolution. Mendel's laws of genetics
This law says that each pair of alternative characteristics behaves independently of each other in a series of generations, as a result of which among the descendants of the first generation (i.e....
This law is called the law of (independent) splitting. Its essence is as follows. When an organism heterozygous for the trait under study develops sex cells - gametes, then one half of them carries one allele of a given gene...
Genetics and evolution. Basic axioms of biology
This law says that each pair of alternative characteristics behaves independently of each other in a series of generations, as a result of which among the descendants of the first generation (i.e....
Law of Radioactive Decay
In the above mathematical expression, it is a decay constant, which characterizes the probability of radioactive decay per unit time and has a dimension of c?1. The minus sign indicates a decrease in the number of radioactive nuclei over time...
Man's place in nature
Phylum Chordata: · the skeleton is formed by the notochord - highly vacuolated connective tissue...
Basic concepts of modern natural science
Certain types of variability are periodic. They are the “vibrations” that determine the levels of being in their hierarchy. Everything in the Universe is alive, not counting the “periphery” - the lower edge of the “hydrogen” scale. Respectively...
Origin and evolution of man
Before talking about the time of the appearance of man, we must clarify the question of the difference between man and animals, since it is the idea of what a man is that forms conclusions about his formation. First, about the similarities between humans and animals...
Correlation between onto- and phylogeny. Law of germinal similarity
Epiphytic lichen biota of mixed and deciduous forests in the vicinity of the village of Zilair, Zilair district
Usually the similarity coefficient is used for this purpose...
In 1828, Karl von Baer formulated a pattern: " The earlier stages of individual development are compared, the more similarities can be found". Comparing the stages of development of embryos of different species and classes of chordates, K. Baer made the following conclusions.
Embryos of animals of the same type are similar in the early stages of development.
They consistently move in their development from more general characteristics of the type to more and more specific ones. Lastly, signs develop that indicate that the embryo belongs to a certain genus, species, and, finally, individual traits.
Embryos of different representatives of the same type gradually become separated from each other.
Baer formulated the laws of germinal similarity:
The most general characters of any large group of animals appear in the embryo earlier than the less general characters;
After the formation of the most general characteristics, less general ones appear, and so on until the appearance of special characteristics characteristic of a given group;
The embryo of any species of animal, as it develops, becomes less and less similar to the embryos of other species and does not pass through the later stages of their development;
The embryo of a highly organized species may resemble the embryo of a more primitive species, but it is never similar to the adult form of this species.
Karl Baer himself did not accept the evolutionary teachings of Charles Darwin, but his laws are considered by biologists as “embryological proof of evolution.”
“For example, during the nine months spent in the womb, a human embryo goes through many stages - from invertebrate to fish, then to amphibian, to reptile, to mammal, to primate, to hominin-like and to man as such.
Von Baer's law implies that evolutionary changes occur more often in later stages of development, while early stages are more evolutionarily conservative. This is because any mutation that affects early development is more likely to produce a significant phenotypic effect than one that affects late development. Because development is continuous and cumulative, changes at an early stage will have increasingly greater consequences than changes at later stages of development. The most likely outcome of any mutation that occurs at an early stage is unfavorable and often fatal. Relatively later (relative to the period of development) mutations are more likely to have no negative effects, and in some cases they can even increase adaptation through subtle changes in the phenotype.( This phenomenon can be illustrated by drawing an analogy with the construction of a skyscraper. If changes are made to the ground floor wall design, there is a high chance that every floor above will be affected, possibly in a negative way. Any changes to the top floor of a skyscraper will not affect the lower floors.)
Von Baer's law is more true for organisms that develop inside the mother (such as mammals) than for species that have a larval stage in which they must fend for themselves.
During intrauterine development, the pressure of natural selection from the external environment leading to change is minimal or absent. However, the larval organism, which independently ensures its survival, is constantly subject to the pressure of natural selection. This explains why the early developmental stages of mammals are so similar across species, but why the larval stage of organisms such as insects is very different from the adult stage.”
At what stages of embryogenesis are the blastocoel, gastrocoel and coelom formed?
Crushing stage. Blastocoel- blastula cavity formed on days 4-5 of cleavage between blastomeres in animal embryos. Filled with a liquid that differs in chemical composition from the environment. The cavity increases the surface area of the embryo, improving its ability to absorb nutrients and oxygen. Reaches its largest size towards the end of cleavage, at the blastula stage. During the process of gastrulation, it is gradually displaced during invagination (invagination) of the embryonic wall or filled (during immigration) with moving cells.
Gastrulation stage. Gastrocoel- the primary gut, the gastrula cavity, which forms in multicellular embryos if gastrulation is carried out by intussusception. The gastrocoel is filled with liquid and communicates with the external environment through a special opening - the blastopore. The walls of the gastrocoel consist of invaginating primary endoderm. Subsequently, the gastrocoel becomes the cavity of the definitive intestine.
Mesoderm Differentiation Stage. Overall- secondary body cavity of multicellular animals. In trochophores, it is formed from specialized mesodermal cells - teloblasts as a result of their division and subsequent formation of cavities within the resulting groups of cells. This method of coelom formation in ontogenesis is called teloblastic. In deuterostomes, the coelom is formed by protrusion of the walls of the primary intestine and separation of the resulting protrusions. This method of coelom formation is called enterocoelous. In both cases, the coelom is considered a mesodermal formation. It differs from the primary body cavity by the presence of its own epithelial lining (wall). The epithelium that forms the lining of the coelom is called coelothelium or mesothelium.
Splitting up. Within 3-4 days, crushing occurs in the oviduct. Light small blastomeres fragment faster and surround dark large ones that remain inside. An embryo without a cavity, consisting of a dense accumulation of blastomeres, is called Morula( formed on the 3-4th day of crushing). On days 4-5, the embryo enters the uterine cavity, from where it absorbs fluid and accumulates it in its cavity - blastocoel . Its wall is formed by small, light blastomeres - trophoblast . Dark cells are pushed to one of the poles and form embryoblast . The resulting blastula called blastocyst , or blastodermic vesicle. Up to 7 days, the blastocyst remains in the uterine cavity in a free state, feeding on the secretion of the uterine glands. This ends the initial (1st week) period of embryogenesis
Intussusception. It is observed in animals with an isolecithal type of eggs (holothuria, lanceolate). The vegetative pole of the blastula invaginates inward. As a result, the opposite poles of the blastoderm practically close together, so that the blastocoel either disappears or a small gap remains. As a result, a two-layer embryo appears, the outer wall of which is the primary ectoderm, and the inner wall is the primary endoderm. The invagination forms the primary intestine-archenteron, or gastrocoel. The opening through which it communicates with the external environment. called the primary mouth, or blastopore.
Mesoderm differentiation. The ventral sections of the mesoderm are not segmented and form a splanchnotome. It is divided into two layers - parietal and visceral, surrounding the secondary body cavity - the coelom.
GASTRULATION
The essence of the gastrulation stage lies in the fact that a single-layer embryo - the blastula - turns into multilayer - two- or three-layer, called gastrula(from Greek gaster - stomach in the diminutive sense).
In primitive chordates, for example, the lancelet, a homogeneous single-layer blastoderm during gastrulation is transformed into an outer germ layer - ectoderm - and an inner germ layer - endoderm. The endoderm forms the primary gut with a cavity inside gastrocele. The hole leading into the gastrocoel is called blastopore or primary mouth. Two germ layers are the defining morphological signs of gastrulation. Their existence at a certain stage of development in all multicellular animals, from coelenterates to higher vertebrates, allows us to think about the homology of the germ layers and the unity of origin of all these animals.
In vertebrates, in addition to the two mentioned during gastrulation, a third germ layer is formed - mesoderm, occupying a place between the ecto- and endoderm. The development of the middle germ layer, which is chordomesoderm, is an evolutionary complication of the gastrulation phase in vertebrates and is associated with the acceleration of their development in the early stages of embryogenesis. In more primitive chordates, such as the lancelet, chordomesoderm is usually formed at the beginning of the next phase after gastrulation - organogenesis. A shift in the time of development of some organs relative to others in descendants compared with ancestral groups is a manifestation heterochrony. Changes in the time of formation of the most important organs in the process of evolution are not uncommon.
The gastrulation process is characterized important cellular transformations, such as directed movements of groups and individual cells, selective proliferation and sorting of cells, the beginning of cytodifferentiation and inductive interactions.
Methods of gastrulation are different. There are four types of spatially directed cell movements that lead to the transformation of the embryo from a single-layer to a multi-layer.
Intussusception - invagination of one of the sections of the blastoderm inward as a whole layer. In the lancelet, the cells of the vegetative pole invaginate; in amphibians, invagination occurs at the border between the animal and vegetative poles in the region of the gray falx. The process of invagination is only possible in eggs with a small or medium amount of yolk.
Epiboly - overgrowth of small cells of the animal pole with larger cells of the vegetative pole that lag behind in the rate of division and are less mobile. This process is clearly expressed in amphibians.
Denomination - separation of blastoderm cells into two layers lying one above the other. Delamination can be observed in the discoblastula of embryos with a partial type of cleavage, such as reptiles, birds, and oviparous mammals. Delamination occurs in the embryoblast of placental mammals, leading to the formation of the hypoblast and epiblast.
Immigration - movement of groups or individual cells that are not united into a single layer. Immigration occurs in all embryos, but is most characteristic of the second phase of gastrulation in higher vertebrates.
In each specific case of embryogenesis, as a rule, several methods of gastrulation are combined.
Features of the gastrulation stage. Gastrulation is characterized by a variety of cellular processes. Mitotic continues cell proliferation, Moreover, it has different intensity in different parts of the embryo. However, the most characteristic feature of gastrulation is movement of cell masses. This leads to a change in the structure of the embryo and its transformation from blastula to gastrula. Happening sorting cells according to their belonging to different germ layers, within which they “recognize” each other.
The gastrulation phase begins cytodifferentiation, which means a transition to the active use of biological information from one’s own genome. One of the regulators of genetic activity is the different chemical composition of the cytoplasm of embryonic cells, established as a result of ovoplasmic segregation. Thus, the ectodermal cells of amphibians are dark in color due to the pigment that entered them from the animal pole of the egg, and the endoderm cells are light, since they originate from the vegetative pole of the egg.
During gastrulation, the role of embryonic induction. It has been shown that the appearance of the primitive streak in birds is the result of an inductive interaction between the hypoblast and the epiblast. The hypoblast is characterized by polarity. A change in the position of the hypoblast relative to the epiblast causes a change in the orientation of the primitive streak.
Zygote formation. Splitting up. Types of crushing.
Zygote- a diploid cell formed as a result of the fusion of haploid female and male gametes (fertilized egg).
Splitting up - continuous mitotic division of diploid cells (zygote and blastomeres) without increasing their total volume.
Zygote formation.
After the fusion of the outer membranes, the sperm nucleus is found inside the egg. From this moment on, the nuclei of the egg and sperm are called the female and male pronuclei. The male pronucleus turns so that the centrioles are closer to the center of the egg and begin to move deeper into the cytoplasm. During this period, DNA doubles in both pronuclei. After this, the sea urchin undergoes fusion of pronuclei and forms a diploid zygote nucleus. In most animals and humans, each pronucleus goes through prophase of mitosis, and their nuclear membranes disintegrate. Then the chromosomal sets of pronuclei combine, and the resulting diploid set forms the metaphase plate. Immediately after this, the first mitosis and cytokinesis occur in the zygote.
Cleavage differs from the normal cell cycle by the absence of G1 and G2 phases and a very short S phase. The interval between crushing divisions is approximately 12-24 hours. The nature of fragmentation and the resulting type of blastula is determined by the number and distribution of yolk inclusions in the cytoplasm of the egg.
Types of crushing:
· Holoblastic crushing type(complete uniform fragmentation) is characteristic of an isolecithal egg. The zygote and blastomeres are completely divided into daughter cells of equal size. With the synchronous type of fragmentation, a geometric progression is observed, 2,4,8,16,32,64,128 blastomeres are sequentially formed; with the asynchronous type, the 3-cell stage is formed.
· Complete uneven crushing occurs in a fertilized telolecithal egg, the egg divides entirely, at the animal pole cleavage occurs faster and numerous small cells are formed, and at the vegetative pole large yolk-laden cells are localized.
· Meroblastic type of crushing (incomplete, partial crushing) occurs in sharply fertilized telolecithal and centrolecithal eggs, the yolk-free cytoplasm is crushed.
· Discoidal incomplete crushing characterized by fragmentation of the cytoplasm containing the nucleus in the area of the animal pole.
By type of symmetry:
· Radial(the axis of the egg cell is the axis of radial symmetry of the dividing blastomeres)
· Spiral(blastomeres resulting from subsequent divisions are located relative to each other in a spiral)
· Bilateral( blastomeres are arranged in such a way that the embryo has only one plane of symmetry)
· Anarchic ( in one species there are different options for the location of blastomeres - chains or shapeless cell mass)
Ontogenesis – implementation of genetic information occurring at all stages.
Ontogenesis is a genetically controlled process. During ontogenesis, the genotype is realized and the phenotype is formed.
Ontogenesis is the individual development of an organism, a set of successive morphological, physiological and biochemical transformations undergone by the organism from the moment of its inception to the end of life. O. includes height, i.e. an increase in body weight, its size, differentiation. The term "O." introduced by E. Haeckel(1866) when he formulated biogenetic law.
The first attempt at a historical substantiation of O. was made by I. f. Meckel. The problem of the relationship between O. and phylogeny was posed by Ch. Darwin and was developed by F. Muller,E. Haeckel and others. All evolutionarily new characters associated with changes in heredity arise in O., but only those that contribute to a better adaptation of the organism to the conditions of existence are preserved in the process natural selection and are passed on to subsequent generations, i.e., they are fixed in evolution. Knowledge of the laws, causes, and factors of oxygen serves as a scientific basis for finding means of influencing the development of plants, animals, and humans, which is of utmost importance for the practice of crop and livestock production, as well as for medicine.
Phylogeny is the historical development of organisms. The term was introduced in German. evolutionist E. Haeckel in 1866. The main task in the study of physiology is to reconstruct the evolutionary transformations of animals, plants, and microorganisms, and to establish on this basis their origin and related relationships between the taxa to which the studied organisms belong. For this purpose, E. Haeckel developed the method of “triple parallelism,” which allows, by comparing data from three sciences—morphology, embryology and paleontology—to reconstruct the course of historical development of the systematic group being studied.
Law of germinal similarity
Explorers of the early 19th century. for the first time they began to pay attention to the similarity of the stages of development of embryos of higher animals with the stages of complexity of organization leading from lowly organized forms to progressive ones. Comparing the stages of development of embryos of different species and classes of chordates, K. Baer made the following conclusions.
1.Embryos of animals of the same type are similar in the early stages of development.
2. They consistently move in their development from more general characteristics of the type to more and more specific ones. Lastly, signs develop that indicate that the embryo belongs to a certain genus, species, and, finally, individual traits.
3. Embryos of different representatives of the same type gradually become separated from each other.
K. Baer, not being an evolutionist, could not connect the patterns of individual development he discovered with the process of phylogenesis. Therefore, the generalizations he made had the meaning of no more than empirical rules.
The subsequent development of the evolutionary idea made it possible to explain the similarity of early embryos by their historical kinship, and their acquisition of more and more particular traits with gradual isolation from each other - the actual isolation of the corresponding classes, orders, families, genera and species in the process of evolution.
Soon after the discovery of the law of embryonic similarity, Charles Darwin showed that this law testifies to the common origin and unity of the initial stages of evolution within the type.
Biogenetic law Haeckel-Müller: every living being in its individual development ( ontogenesis) repeats to a certain extent the forms traversed by his ancestors or his species ( phylogenesis).
Ontogenesis - repetition of phylogeny
Comparing the ontogenesis of crustaceans with the morphology of their extinct ancestors, F. Muller concluded that living crustaceans in their development repeat the path traversed by their ancestors. The transformation of ontogeny in evolution, according to F. Muller, is carried out due to its lengthening by adding additional stages or extensions to it. Based on these observations, as well as studying the development of chordates, E. Haeckel (1866) formulated the basic biogenetic law, according to which Ontogenesis is a short and rapid repetition of phylogeny.
The repetition of structures characteristic of ancestors in the embryogenesis of descendants is called recapitulations. Not only the morphological characteristics of the notochord, the anlage of the gill slits and gill arches in all chordates recapitulate, but also the features of the biochemical organization and physiology. Thus, in the evolution of vertebrates, there is a gradual loss of enzymes necessary for the breakdown of uric acid, a product of purine metabolism. In most invertebrates, the final product of the breakdown of uric acid is ammonia, in amphibians and fish it is urea, in many reptiles it is allantoin, and in some mammals uric acid is not broken down at all and is excreted in the urine. In the embryogenesis of mammals and humans, biochemical and physiological recapitulations are noted: the release of ammonia by early embryos, later urea, then allantoin, and in the last stages of development - uric acid.
However, in the ontogenesis of highly organized organisms, a strict repetition of the stages of historical development is not always observed, as follows from the biogenetic law. Thus, the human embryo never repeats the adult stages of fish, amphibians, reptiles and mammals, but is similar in a number of features only to their embryos. The early stages of development remain the most conservative, due to which they recapitulate more completely than the later ones. This is due to the fact that one of the most important mechanisms for integrating the early stages of embryogenesis is embryonic induction, and the structures of the embryo that form first, such as the notochord, neural tube, pharynx, gut and somites, represent the organizational centers of the embryo, from which depends on the entire course of development.
The genetic basis of recapitulation lies in the unity of the mechanisms of genetic control of development, which is preserved on the basis of common genes for the regulation of ontogenesis, which are inherited by related groups of organisms from common ancestors.
Recapitulation(from Latin recapitulatio - repetition) - a concept used in biology to denote the repetition in individual development of characteristics characteristic of an earlier stage of evolutionary development.
Ontogenesis as the basis of phylogeny. Cenogenesis. Autonomy of ontogeny. Philembryogenesis. The doctrine of A.N. Severtsov about phylembryogenesis. Mechanisms of their occurrence. Heterochronies and heterotopies of biological structures in the evolution of ontogenesis.
Relying only on the basic biogenetic law, it is impossible to explain the process of evolution: the endless repetition of what has been passed does not in itself give birth to something new. Since life exists on Earth thanks to the change of generations of specific organisms, its evolution occurs due to changes occurring in their ontogenies. These changes boil down to the fact that specific ontogenies deviate from the path laid out by ancestral forms and acquire new features.
Such deviations include, for example, cenogenesis - adaptations that arise in embryos or larvae and adapt them to the characteristics of their environment. In adult organisms, cenogenesis is not preserved. Examples of coenogenesis are horny formations in the mouth of the larvae of tailless amphibians, which make it easier for them to feed on plant foods. During the process of metamorphosis in the frog, they disappear and the digestive system is rebuilt to feed on insects and worms. Coenogenesis in amniotes includes the embryonic membranes, yolk sac and allantois, and in placental mammals and humans also the placenta and umbilical cord.
Cenogenesis, appearing only in the early stages of ontogenesis, does not change the type of organization of the adult organism, but provides a higher probability of survival of the offspring. They may be accompanied by a decrease in fertility and a lengthening of the embryonic or larval period, due to which the organism in the postembryonic or postlarval period of development turns out to be more mature and active. Having arisen and proven useful, cenogeneses will be reproduced in subsequent generations. Thus, the amnion, which first appeared in the ancestors of reptiles in the Carboniferous period of the Paleozoic era, is reproduced in all vertebrates developing on land, both in oviparous reptiles and birds, and in placental mammals.
Another type of phylogenetically significant transformations of phylogeny is phylembryogenesis. They represent deviations from ontogenesis characteristic of ancestors, manifested in embryogenesis, but having adaptive significance in adult forms. Thus, hair buds appear in mammals at very early stages of embryonic development, but the hair itself is important only in adult organisms.
Such changes in ontogenesis, being useful, are fixed by natural selection and reproduced in subsequent generations. These changes are based on the same mechanisms that cause congenital malformations: disruption of cell proliferation, their movement, adhesion, death or differentiation (see § 8.2 and 9.3). However, just like cenogenesis, they are distinguished from defects by adaptive value, i.e. usefulness and fixation by natural selection in phylogenesis.
Depending on the stages of embryogenesis and morphogenesis of specific structures, developmental changes that have the significance of phylembryogenesis occur, three types are distinguished.
1.Anabolism, or extensions, arise after the organ has practically completed its development, and are expressed in the addition of additional stages that change the final result.
Anabolisms include such phenomena as the acquisition of a specific body shape by flounder only after the egg hatches into a fry, indistinguishable from other fish, as well as the appearance of curvatures of the spine, fusion of sutures in the brain skull, and the final redistribution of blood vessels in the body of mammals and humans.
2.Deviations - deviations that arise in the process of organ morphogenesis. An example is the development of the heart in the ontogenesis of mammals, in which it recapitulates the tube stage, two-chamber and three-chamber structure, but the stage of formation of an incomplete septum, characteristic of reptiles, is replaced by the development of a septum, constructed and located differently and characteristic only of mammals (see § 14.4) .The development of the lungs in mammals also reveals recapitulation of the early stages of the ancestors; later morphogenesis proceeds in a new way (see section 14.3.4).
Rice. 13.9.Transformations of onto- and phylogeny in connection with emerging phylembryogenesis
Letters indicate stages of ontogenesis, numbers indicate phylembryogenetic transformations
3.Archallaxis - changes that are detected at the level of primordia and are expressed in a violation of their division, early differentiation, or in the appearance of fundamentally new primordia. A classic example of archallaxis is
the development of hair in mammals, the anlage of which occurs at very early stages of development and from the very beginning differs from the anlage of other vertebrate skin appendages (see § 14.1).
According to the type of archallaxis, the notochord appears in primitive skullless fish, the cartilaginous spine in cartilaginous fish (see Section 14.2.1.1), and the nephrons of the secondary kidney develop in reptiles (see Section 14.5.1).
It is clear that during evolution due to anabolism in the ontogenies of descendants, the basic biogenetic law is fully realized, i.e. recapitulations of all ancestral stages of development occur. In case of deviations, early ancestral stages recapitulate, and later ones are replaced by development in a new direction. Archallaxis completely prevent recapitulation in the development of these structures, changing their very rudiments.
If we compare the diagram of phylembryogenesis with K. Baer’s table (Fig. 13.9), illustrating the law of germinal similarity, it becomes clear that Baer was already very close to the discovery of phylembryogenesis, but the absence of an evolutionary idea in his reasoning did not allow him to be more than 100 years ahead of scientific thought .
In the evolution of ontogenesis, anabolism most often occurs as phylembryogenesis, which only to a small extent changes the entire development process. Deviations as violations of the morphogenetic process in embryogenesis are often rejected by natural selection and are therefore much less common. Archallaxis occur most rarely in evolution due to the fact that they change the entire course of embryogenesis, and if such changes affect the rudiments of vital organs or organs that are embryonic organizational centers (see Section 8.2.6), then they often turn out to be incompatible with life.
In the same phylogenetic group, evolution in different organ systems can occur due to different phylembryogenesis.
Thus, in the ontogenesis of mammals, all stages of the development of the axial skeleton in the vertebrate subtype are traced (anabolism), in the development of the heart only early stages recapitulate (deviation), and in the development of skin appendages there is no recapitulation at all (archallaxis). Knowledge of the types of phylembryogenesis in the evolution of organ systems of chordates is necessary for the doctor to predict the possibility of the occurrence of congenital malformations of an atavistic nature in fetuses and newborns (see Section 13.3.4). Indeed, if atavistic malformations are possible in an organ system evolving through anabolism and deviations due to the recapitulation of ancestral states, then in the case of archallaxis this is completely excluded.
In addition to cenogenesis and phylembryogenesis, deviations in the time of organ formation can also be found in the evolution of ontogenesis - heterochrony - and places of their development - heterotopia. Both the first and second lead to changes in the mutual correspondence of developing structures and undergo strict control of natural selection. Only those heterochronies and heterotopies that prove useful are preserved. Examples of such adaptive heterochronies are shifts in the time of the formation of the most vital organs in groups evolving according to the type of arogenesis. Thus, in mammals, and especially in humans, the differentiation of the forebrain significantly advances the development of its other parts.
Heterotopies lead to the formation of new spatial and functional connections between organs, ensuring their further joint evolution. Thus, the heart, located under the pharynx of fish, ensures efficient flow of blood into the gill arteries for gas exchange. Moving to the retrosternal region in terrestrial vertebrates, it develops and functions in a single complex with the new respiratory organs - the lungs, and here, first of all, performs the function of delivering blood to the respiratory system for gas exchange.
Heterochronies and heterotopies, depending on at what stages of embryogenesis and organ morphogenesis they appear, can be regarded as phylembryogenesis of different types. Thus, the movement of the rudiments of the brain, leading to its bending, characteristic of amniotes, and manifested at the initial stages of its differentiation, is archallaxis, and heterotopia of the testis in humans from the abdominal cavity through the inguinal canal to the scrotum, observed at the end of embryogenesis after its final formation, - typical anabolia.
Sometimes processes of heterotopy, identical in results, can be phylembryogenesis of different types. For example, in various classes of vertebrates, movement of the limb girdles is very common. In many groups of fish leading a bottom-dwelling lifestyle, the ventral fins (hind limbs) are located anterior to the pectoral fins, and in mammals and humans, the shoulder girdle and forelimbs in the definitive state are located significantly caudal to the place of their initial formation. In this regard, the innervation of the shoulder girdle in them is carried out by nerves connected not with the thoracic, but with the cervical segments of the spinal cord. In the fish mentioned above, the pelvic fins are innervated not by the posterior trunk nerves, but by the anterior segments, located anterior to the centers of innervation of the pectoral fins. This indicates heterotopia in the formation of fins already at the stage of the earliest rudiments, while the movement of the anterior girdle of the limbs in humans occurs at later stages, when their innervation has already been fully accomplished. Obviously, in the first case, heterotopia represents archallaxis, while in the second, anabolism.
Cenogenesis, phylembryogenesis, as well as heterotopia and heterochrony, having proven useful, are fixed in the offspring and reproduced in subsequent generations until new adaptive changes in ontogenesis displace them and replace them. Thanks to this, ontogenesis not only briefly repeats the evolutionary path traversed by our ancestors, but also paves new directions for phylogeny in the future.
Cenogenesis
(from the Greek kainós - new and ... genesis (See ... genesis)
an adaptation of an organism that occurs at the embryonic (fetal) or larval stage and does not persist in the adult. Examples of c. are the mammalian placenta, which provides the fetus with breathing, nutrition, and excretion; external gills of amphibian larvae; egg tooth in birds, used by chicks to break through the egg shell; organs of attachment in ascidian larvae, swimming tail in trematode larvae - cercariae, etc. The term “C.” introduced in 1866 by E. Haeckel to designate those characteristics that, disrupting the manifestations of palingenesis (See. Palingenesis), i.e., repetitions of distant stages of phylogenesis in the process of embryonic development of an individual, do not allow us to trace the sequence of stages of the phylogeny of their ancestors during the ontogeny of modern forms, i.e., they violate Biogenetic law. At the end of the 19th century. Any change in the course of ontogenesis characteristic of the ancestors began to be called color (German scientists E. Mehnert, F. Keibel, and others). Modern understanding of the term “C.” was formed as a result of the work of A. N. Severtsov, who retained for this concept only the meaning of provisional devices, or embryo-adaptations. see also Philembryogenesis.
Cenogenesis(Greek kainos new + genesis origin, formation) - the appearance in the embryo or larva of adaptations to living conditions that are not characteristic of the adult stages, for example. formation of membranes in the embryos of higher animals.
Philembryogenesis
(from Greek phýlon - tribe, genus, species and Embryogenesis
PHYLEMBRYOGENESIS (from the Greek phylon - genus, tribe, embryon - embryo and genesis - origin), evolutionary change ontogeny organs, tissues and cells, associated with both progressive development and reduction. The doctrine of phylembryogenesis was developed by a Russian evolutionary biologist A.N. Severtsov. The modes (methods) of phylembryogenesis differ in the time of occurrence during the development of these structures.
If the development of a certain organ in descendants continues after the stage at which it ended in the ancestors, anabolia occurs (from the Greek anabole - rise) - an extension of the final stage of development. An example is the formation of a four-chambered heart in mammals. Amphibians have a three-chambered heart: two atria and one ventricle. In reptiles, a septum develops in the ventricle (first anabolia), but in most of them this septum is incomplete - it only reduces the mixing of arterial and venous blood. In crocodiles and mammals, the development of the septum continues until the complete separation of the right and left ventricles (second anabolia). In children, sometimes, as an atavism, the interventricular septum is underdeveloped, which leads to a serious illness requiring surgical intervention.
Prolonging the development of an organ does not require profound changes in the previous stages of its ontogenesis, therefore anabolism is the most common method of phylembryogenesis. The stages of organ development preceding anabolism remain comparable to the stages phylogeny ancestors (i.e. they are recapitulations) and can serve for its reconstruction (see. Biogenetic law). If the development of an organ at intermediate stages deviates from the path along which its ontogenesis took place in its ancestors, deviation occurs (from Late Latin deviatio - deviation). For example, in fish and reptiles, scales appear as thickenings of the epidermis and the underlying connective tissue layer of the skin - the corium. Gradually thickening, this anlage bends outward. Then in fish the corium ossifies, the forming bone scales pierce the epidermis and move to the surface of the body. In reptiles, on the contrary, bone is not formed, but the epidermis becomes keratinized, forming the horny scales of lizards and snakes. In crocodiles, the corium can ossify, forming the bony basis of the horny scales. Deviations lead to a more profound restructuring of ontogenesis than anabolism, so they are less common.
Changes in the primary rudiments of organs - archallaxis (from the Greek arche - beginning and allaxis - change) occur least often. In case of deviation, recapitulation can be traced from the origin of the organ to the moment of developmental deviation. In archallaxis there is no recapitulation. An example is the development of vertebral bodies in amphibians. In fossil amphibians - stegocephalians and in modern tailless amphibians, the vertebral bodies are formed around a chord of several, usually three on each side of the body, separate anlage, which then merge to form the vertebral body. In tailed amphibians these anlages do not appear. Ossification grows above and below, covering the notochord, so that a bone tube is immediately formed, which, thickening, becomes the vertebral body. This archallaxis is the reason for the still debated question of the origin of tailed amphibians. Some scientists believe that they descended directly from lobe-finned fish, regardless of other land vertebrates. Others say that tailed amphibians diverged very early from other amphibians. Still others, neglecting the development of the vertebrae, prove the close relationship of tailed and tailless amphibians.
Organ reduction, which have lost their adaptive significance, also occurs through phylembryogenesis, mainly through negative anabolism - loss of the final stages of development. In this case, the organ either underdevelops and becomes a vestige, or undergoes reverse development and completely disappears. An example of a rudiment is the human appendix - an underdeveloped cecum; an example of complete disappearance is the tail of frog tadpoles. Throughout its life in water, the tail grows, new vertebrae and muscle segments are added at its end. During metamorphosis, when the tadpole turns into a frog, the tail dissolves, and the process occurs in the reverse order - from the end to the base. Phylembryogenesis is the main method of adaptive changes in the structure of organisms during phylogenesis.
Principles (methods) of phylogenetic transformations of organs and functions. Correspondence between structure and function in living systems. Multifunctionality. Quantitative and qualitative changes in the functions of biological structures.
GENERAL REGULARITIES
EVOLUTION OF ORGANS
An organism, or individual, is a separate living being that, in the process of ontogenesis, exhibits all the properties of a living thing. The constant interaction of an individual with the environment in the form of organized flows of energy and matter maintains its integrity and development. Structurally, the body is an integrated hierarchical system built from cells, tissues, organs and systems that ensure its vital functions. Let's take a closer look at the organs and life support systems.
Authority call a historically developed specialized system of tissues, characterized by delimitation, constancy of shape, localization, internal structure of circulatory and innervation pathways, development in ontogenesis and specific functions. The structure of organs is often very complex. Most of them are multifunctional, i.e. performs several functions simultaneously. At the same time, various organs may participate in the implementation of any complex function.
A group of organs of similar origin that combine to perform a complex function is called system(circulatory, excretory, etc.).
If a group of organs of different origin performs the same function, it is called apparatus. An example is the respiratory apparatus, which consists of both the respiratory organs themselves and the elements of the skeleton and muscular system that provide respiratory movements.
In the process of ontogenesis, development and often replacement of some organs by others occurs. The organs of a mature organism are called definitive; organs that develop and function only in embryonic or larval development - provisional. Examples of provisional organs are the gills of amphibian larvae, the primary kidney, and the embryonic membranes of higher vertebrates (amniotes).
In historical development, organ transformations can be progressive or regressive. In the first case, the organs increase in size and become more complex in their structure; in the second, they decrease in size and their structure becomes simpler.
If two organisms located at different levels of organization have organs that are built according to a single plan, located in the same place and develop in a similar way from the same embryonic rudiments, then this indicates the relationship of these organisms. Such organs are called homologous. Homologous organs often perform the same function (for example, the heart of fish, amphibians, reptiles and mammals), but in the process of evolution the functions can change (for example, the forelimbs of fish and amphibians, reptiles and birds).
When unrelated organisms live in the same environments, they may develop similar adaptations, which manifest themselves in the appearance similar organs. Similar organs perform the same functions, but their structure, location and development are sharply different. Examples of such organs are the wings of insects and birds, the limbs and jaw apparatus of arthropods and vertebrates.
The structure of organs strictly corresponds to the functions they perform. Moreover, in the historical transformations of organs, a change in functions is certainly accompanied by a change in the morphological characteristics of the organ.