Ko je formulisao zakon germinativne sličnosti. Individualni i istorijski razvoj. Zakon germinativne sličnosti. Biogenetski zakon. Rekapitulacija. Biogenetski zakon i embrionalna varijabilnost
Ontogeneza – implementacija genetske informacije koja se javlja u svim fazama.
Ontogeneza je genetski kontrolisan proces. Tokom ontogeneze ostvaruje se genotip i formira fenotip.
Ontogeneza je individualni razvoj organizma, skup uzastopnih morfoloških, fizioloških i biohemijskih transformacija kroz koje organizam prolazi od trenutka svog nastanka do kraja života. O. uključuje rast, odnosno povećanje tjelesne težine, njegove veličine i diferencijaciju. Izraz "O." uveo E. Haeckel (1866) kada je formulisao biogenetski zakon.
Prvi pokušaj istorijskog potkrepljenja O. napravio je I. f. Meckel. Problem odnosa između O. i filogenije postavio je C. Darwin, a razvio ga F. Müller, E. Haeckel i dr. Sve nove evolucijske osobine povezane s promjenama u nasljeđu nastaju kod O., ali samo one koje doprinose boljoj adaptaciji organizma na uslove života se čuvaju u procesu prirodne selekcije i prenose na sljedeće generacije, tj. fiksirani su u evoluciji. Poznavanje zakona, uzroka i faktora kiseonika služi kao naučna osnova za pronalaženje sredstava uticaja na razvoj biljaka, životinja i ljudi, što je od izuzetne važnosti za biljnu i stočarsku proizvodnju, kao i za medicinu. .
Filogenija je istorijski razvoj organizama. Termin je uveden u njemački jezik. evolucionista E. Haeckel 1866. Glavni zadatak u proučavanju fiziologije je rekonstruisati evolucionu transformaciju životinja, biljaka i mikroorganizama i na osnovu toga utvrditi njihovo porijeklo i srodne odnose između svojti kojima proučavani organizmi pripadaju. U tu svrhu, E. Haeckel je razvio metodu „trostrukog paralelizma“, koja omogućava da se upoređivanjem podataka iz tri nauke — morfologije, embriologije i paleontologije — rekonstruiše tok istorijskog razvoja sistematske grupe koja se proučava.
Zakon germinativne sličnosti
Istraživači ranog 19. veka. po prvi put su počeli obraćati pažnju na sličnost faza razvoja embriona viših životinja sa fazama složenosti organizacije koje vode od nisko organiziranih oblika ka progresivnim. Upoređujući faze razvoja embriona različitih vrsta i klasa hordata, K. Baer je došao do sljedećih zaključaka.
1. Embrioni životinja istog tipa su slični u ranim fazama razvoja.
2. Dosljedno se kreću u svom razvoju od opštijih karakteristika tipa ka sve specifičnijim. Konačno, razvijaju se znakovi koji ukazuju da embrij pripada određenom rodu, vrsti i, konačno, individualnim osobinama.
3. Embrioni različitih predstavnika istog tipa postepeno se odvajaju jedan od drugog.
K. Baer, budući da nije evolucionista, nije mogao povezati obrasce individualnog razvoja koje je otkrio s procesom filogeneze. Stoga su generalizacije koje je napravio imale značenje samo empirijskih pravila.
Naknadni razvoj evolucione ideje omogućio je da se sličnost ranih embriona objasni njihovim istorijskim srodstvom, i njihovo sticanje sve specifičnijih osobina uz postepenu izolaciju jedna od druge - stvarna izolacija odgovarajućih klasa, redova, porodica, rodova i vrsta u procesu evolucije.
Ubrzo nakon otkrića zakona embrionalne sličnosti, Charles Darwin je pokazao da ovaj zakon svjedoči o zajedničkom porijeklu i jedinstvu početnih faza evolucije unutar tipa.
Biogenetski zakon Haeckel-Müller: svako živo biće u svom individualnom razvoju (ontogenezi) u određenoj mjeri ponavlja oblike koje su prošli njegovi preci ili njegove vrste (filogeneza).
Ontogeneza - ponavljanje filogeneze
Uspoređujući ontogenezu rakova sa morfologijom njihovih izumrlih predaka, F. Muller je zaključio da živi rakovi u svom razvoju ponavljaju put koji su prošli njihovi preci. Transformacija ontogeneze u evoluciji, prema F. Mulleru, vrši se zbog njenog produžavanja dodavanjem dodatnih faza ili proširenja. Na osnovu ovih zapažanja, kao i proučavanja razvoja hordata, E. Haeckel (1866) je formulisao osnovni biogenetički zakon, prema kojem je Ontogeneza je kratko i brzo ponavljanje filogenije.
Ponavljanje struktura karakterističnih za pretke u embriogenezi potomaka naziva se rekapitulacije. Rekapitulirajte ne samo morfološke karakteristike - notohordu, škržne proreze i škržne lukove kod svih hordata, već i karakteristike biohemijske organizacije i fiziologije. Dakle, u evoluciji kralježnjaka dolazi do postepenog gubitka enzima neophodnih za razgradnju mokraćne kiseline, produkta metabolizma purina. Kod većine beskičmenjaka krajnji produkt razgradnje mokraćne kiseline je amonijak, kod vodozemaca i riba to je urea, kod mnogih gmizavaca alantoin, a kod nekih sisara mokraćna kiselina se uopće ne razgrađuje i izlučuje se urinom. U embriogenezi sisara i ljudi bilježe se biohemijske i fiziološke rekapitulacije: oslobađanje amonijaka od ranih embrija, kasnije uree, zatim alantoina, a u posljednjim fazama razvoja - mokraćne kiseline.
Međutim, u ontogenezi visokoorganiziranih organizama ne opaža se uvijek striktno ponavljanje faza historijskog razvoja, kao što slijedi iz biogenetskog zakona. Dakle, ljudski embrion nikada ne ponavlja odrasle stadijume riba, vodozemaca, gmizavaca i sisara, već je po nizu osobina sličan samo njihovim embrionima. Rane faze razvoja ostaju najkonzervativnije, zbog čega se potpunije rekapituliraju od kasnijih. To je zbog činjenice da je jedan od najvažnijih mehanizama za integraciju ranih faza embriogeneze embrionalna indukcija, a strukture embrija koje se prve formiraju, kao što su notohorda, neuralna cijev, ždrijelo, crijeva i somiti, predstavljaju organizacione centre embrija, od kojih zavisi ceo tok razvoja.
Genetska osnova rekapitulacije leži u jedinstvu mehanizama genetičke kontrole razvoja, koji se čuva na osnovu zajedničkih gena za regulaciju ontogeneze, koje nasljeđuju srodne grupe organizama od zajedničkih predaka.
Rekapitulacija(od latinskog recapitulatio - ponavljanje) - koncept koji se koristi u biologiji za označavanje ponavljanja u individualnom razvoju karakteristika karakterističnih za raniju fazu evolucijskog razvoja.
Ulaznica 96.
Ontogeneza kao osnova filogeneze. Cenogeneza. Autonomija ontogenije. Filembriogeneza. Doktrina A.N. Severtsova o filebriogenezi. Mehanizmi njihovog nastanka. Heterohronije i heterotopije bioloških struktura u evoluciji ontogeneze.
Oslanjajući se samo na osnovni biogenetički zakon, nemoguće je objasniti proces evolucije: beskonačno ponavljanje onoga što je prošlo samo po sebi ne rađa nešto novo. Budući da život na Zemlji postoji zahvaljujući smjeni generacija određenih organizama, njegova evolucija nastaje zbog promjena koje se dešavaju u njihovoj ontogeniji. Ove promjene se svode na činjenicu da specifične ontogenije odstupaju od puta koji su zacrtali oblici predaka i dobijaju nove karakteristike.
Takva odstupanja uključuju npr. cenogeneza - adaptacije koje nastaju u embrionima ili larvama i prilagođavaju ih karakteristikama njihovog okruženja. Kod odraslih organizama cenogeneza nije očuvana. Primjeri koenogeneze su rožnate formacije u ustima ličinki vodozemaca bez repa, koje im olakšavaju hranu biljnom hranom. Tokom procesa metamorfoze u žabi, oni nestaju i probavni sistem se obnavlja da se hrani insektima i crvima. Koenogeneza kod amniota uključuje embrionalne membrane, žumančanu vreću i alantois, a kod placentnih sisara i ljudi također uključuje posteljicu i pupčanu vrpcu.
Cenogeneza, koja se pojavljuje tek u ranim fazama ontogeneze, ne mijenja tip organizacije odraslog organizma, ali daje veću vjerovatnoću preživljavanja potomstva. Oni mogu biti praćeni smanjenjem plodnosti i produženjem embrionalnog ili larvalnog perioda, zbog čega se organizam u postembrionalnom ili postlarvalnom razdoblju razvoja pokazuje zrelijim i aktivnijim. Nakon što su nastale i pokazale se korisnim, cenogeneze će se razmnožavati u narednim generacijama. Dakle, amnion, koji se prvi put pojavio kod predaka gmizavaca u karbonskom periodu paleozojske ere, reproducira se kod svih kralježnjaka koji se razvijaju na kopnu, kako kod jajonosnih reptila i ptica, tako i kod placentnih sisara.
Druga vrsta filogenetski značajnih transformacija filogenije je filebriogeneza. Predstavljaju odstupanja od ontogeneze karakteristična za pretke, koja se manifestuju u embriogenezi, ali imaju adaptivni značaj kod odraslih oblika. Tako se pupoljci dlake pojavljuju kod sisara u vrlo ranim fazama embrionalnog razvoja, ali sama dlaka je važna samo kod odraslih organizama.
Takve promjene u ontogenezi, budući da su korisne, fiksiraju se prirodnom selekcijom i reprodukuju u narednim generacijama. Ove promjene se temelje na istim mehanizmima koji uzrokuju kongenitalne malformacije: poremećena proliferacija ćelija, kretanje, adhezija, smrt ili diferencijacija. Međutim, kao i cenogeneza, od poroka se razlikuju po adaptivnoj vrijednosti, tj. korisnost i fiksacija prirodnom selekcijom u filogenezi.
U zavisnosti od faza embriogeneze i morfogeneze određenih struktura, nastaju razvojne promene koje imaju značaj filebriogeneze, razlikuju se tri tipa.
1. anabolizam, ili ekstenzije, nastaju nakon što je organ praktično završio svoj razvoj, a izražavaju se u dodavanju dodatnih faza koje mijenjaju konačni rezultat.
Anabolizmi uključuju fenomene kao što je iverak sticanje specifičnog oblika tijela tek nakon što se iz jajeta izleže mladica, koja se ne razlikuje od ostalih riba, kao i pojavu zakrivljenosti kralježnice, spajanje šavova u moždanoj lubanji i konačno preraspodjela krvnih žila u tijelu sisara i ljudi.
2. odstupanja - devijacije koje nastaju u procesu morfogeneze organa. Primjer je razvoj srca u ontogenezi sisara, u kojem ono rekapitulira stadij cijevi, dvokomornu i trokomornu strukturu, ali fazu formiranja nepotpunog septuma, karakterističnog za gmizavce, zamjenjuje razvojem pregrade, građene i locirane drugačije i karakteristične samo za sisare. U razvoju pluća kod sisara takođe se nalazi rekapitulacija ranih faza predaka, a kasnija morfogeneza teče na nov način.
3. arhalaksija - promjene koje se otkrivaju na nivou primordija i izražavaju se u kršenju njihove podjele, ranoj diferencijaciji ili u pojavi fundamentalno novih primordija. Klasičan primjer arhalaksije je
razvoj dlake kod sisara, čije se zarastanje javlja u vrlo ranim fazama razvoja i od samog početka razlikuje se od dlaka kod drugih kožnih dodataka kralježnjaka.
Prema vrsti arhalaksije, notohorda se pojavljuje kod primitivnih riba bez lubanje, hrskavična kralježnica kod hrskavičnih riba, a nefroni sekundarnog bubrega razvijaju se kod gmizavaca.
Jasno je da se tokom evolucije zbog anabolizma u ontogenijima potomaka u potpunosti ostvaruje osnovni biogenetski zakon, tj. dešavaju se rekapitulacije svih faza razvoja predaka. U slučaju odstupanja, rani stadijumi predaka se rekapituliraju, a kasnije se zamjenjuju razvojem u novom smjeru. Arhalaksija u potpunosti sprečava rekapitulaciju u razvoju ovih struktura, mijenjajući same njihove rudimente.
Ako uporedimo dijagram filebriogeneze sa tabelom K. Baera, koja ilustruje zakon germinalne sličnosti, postaje jasno da je Baer već bio vrlo blizu otkrića filebriogeneze, ali je odsustvo evolucijske ideje u njegovom rasuđivanju učinilo ne dozvoli mu da bude više od 100 godina ispred naučne misli.
U evoluciji ontogeneze anabolizam se najčešće javlja kao filebriogeneza, koja samo u maloj mjeri mijenja cjelokupni razvojni proces. Odstupanja kao kršenja morfogenetskog procesa u embriogenezi često se odbacuju prirodnom selekcijom i stoga su mnogo rjeđa. Arhalaksije se najrjeđe javljaju u evoluciji zbog činjenice da mijenjaju cijeli tok embriogeneze, a ako takve promjene zahvaćaju rudimente vitalnih organa ili organa koji su važni embrionalni organizacijski centri, onda se često ispostavi da su nespojive sa životom.
U istoj filogenetskoj grupi, evolucija u različitim sistemima organa može se dogoditi zbog različite filebriogeneze.
Tako se u ontogenezi sisara prate svi stadijumi razvoja aksijalnog skeleta u podtipu kičmenjaka (anabolizam), u razvoju srca se rekapituliraju samo rani stadijumi (devijacija), a u razvoju kožnih privjesaka postoji nema rekapitulacije uopšte (arhalaksija). Poznavanje tipova filebriogeneze u evoluciji organskih sistema hordata je neophodno da bi ljekar mogao predvidjeti mogućnost pojave kongenitalnih malformacija atavističke prirode kod fetusa i novorođenčadi. Zaista, ako su u organskom sistemu koji se razvija anabolizmom i devijacijom, mogući atavistički defekti u razvoju zbog rekapitulacije stanja predaka, onda je to u slučaju arhalaksije potpuno isključeno.
Osim cenogeneze i filebriogeneze, odstupanja u vremenu formiranja organa mogu se naći i u evoluciji ontogeneze - heterohronija - i mesta njihovog razvoja - heterotopija. I prvi i drugi dovode do promjena u međusobnoj korespondenciji struktura u razvoju i podliježu strogoj kontroli prirodne selekcije. Sačuvane su samo one heterohronije i heterotopije koje se pokažu korisnima. Primjeri takvih adaptivnih heterohronija su pomaci u vremenu formiranja najvitalnijih organa u grupama koje se razvijaju prema tipu arogeneze. Dakle, kod sisara, a posebno kod ljudi, diferencijacija prednjeg mozga značajno pospješuje razvoj ostalih njegovih dijelova.
Heterotopije dovode do stvaranja novih prostornih i funkcionalnih veza između organa, osiguravajući njihovu daljnju zajedničku evoluciju. Tako srce, koje se nalazi ispod ždrijela ribe, osigurava efikasan protok krvi u škržne arterije za razmjenu plinova. Prelazeći u retrosternalnu regiju kod kopnenih kralježnjaka, razvija se i funkcionira u jednom kompleksu s novim respiratornim organima - plućima, obavljajući ovdje prvenstveno funkciju isporuke krvi u respiratorni sistem za izmjenu plinova.
Heterohronije i heterotopije, u zavisnosti od toga u kojim fazama embriogeneze i morfogeneze organa se pojavljuju, mogu se smatrati filebriogenezom različitih tipova. Dakle, kretanje rudimenata mozga, koje dovodi do njegovog savijanja, karakteristično za amniote, a koje se manifestuje u početnim fazama njegove diferencijacije, je arhalaksija i heterotopija testisa kod ljudi od trbušne šupljine kroz ingvinalni kanal do skrotum, uočen na kraju embriogeneze nakon njegovog konačnog formiranja, - tipična anabolija.
Ponekad procesi heterotopije, identični u rezultatima, mogu biti filebriogeneza različitih tipova. Na primjer, u različitim klasama kralježnjaka, pomicanje pojasa udova je vrlo uobičajeno. U mnogim grupama riba koje vode način života na dnu, trbušna peraja (zadnji udovi) nalaze se ispred prsnih peraja, a kod sisara i ljudi rameni pojas i prednji udovi u konačnom stanju nalaze se znatno kaudalno od mjesta njihovo početno formiranje. S tim u vezi, inervaciju ramenog pojasa u njima provode živci povezani ne s torakalnim, već s cervikalnim segmentima kičmene moždine. Kod gore navedenih riba, karlične peraje inerviraju ne zadnji nervi trupa, već prednji segmenti, koji se nalaze ispred centara inervacije prsnih peraja. To ukazuje na heterotopiju u formiranju peraja već u fazi najranijih rudimenata, dok se pomicanje prednjeg pojasa udova kod ljudi javlja u kasnijim fazama, kada je njihova inervacija već u potpunosti ostvarena. Očigledno, u prvom slučaju heterotopija predstavlja arhalaksu, au drugom anaboliju.
Cenogeneza, filebriogeneza, kao i heterotopija i heterohronija, koji su se pokazali korisnima, fiksiraju se u potomstvu i reprodukuju u narednim generacijama sve dok ih nove adaptivne promjene u ontogenezi ne istisnu i zamijene. Zahvaljujući tome, ontogeneza ne samo da nakratko ponavlja evolucijski put koji su prošli naši preci, već i utire nove pravce filogeneze u budućnosti.
Cenogeneza(od grčkog kainós - novi i ... geneza (Vidi ... geneza) adaptacija organizma koja se javlja u embrionalnom (fetalnom) ili larvalnom stadiju i ne perzistira u odrasloj jedinki. Primeri c. su placenta sisara, koja fetusu obezbeđuje disanje, ishranu i izlučivanje; vanjske škrge larvi vodozemaca; zub jajeta kod ptica, koji pilići koriste da probiju ljusku jajeta; organi vezivanja u larvi ascidijana, plivajući rep u larvi trematoda - cercariae, itd. Izraz "C." koju je 1866. uveo E. Haeckel da označi one karakteristike koje, narušavajući manifestacije palingeneze , odnosno ponavljanja udaljenih faza filogeneze u procesu embrionalnog razvoja pojedinca, ne dozvoljavaju nam da pratimo slijed faza filogeneze njihovih predaka tokom ontogeneze modernih oblika, odnosno krše biogenetski zakon. Krajem 19. vijeka. Svaka promjena u toku ontogeneze karakteristična za pretke počela je da se naziva bojom (njemački naučnici E. Mehnert, F. Keibel i drugi). Moderno razumijevanje pojma "C." nastao je kao rezultat rada A. N. Severtsova, koji je za ovaj koncept zadržao samo značenje privremenih uređaja, odnosno embrionalnih adaptacija. Vidi također Filembriogeneza.
Cenogeneza(grč. kainos novi + geneza porijeklo, formiranje) - pojava u embrionu ili larvi adaptacija na uslove života koji nisu karakteristični za odrasle faze, na primjer. formiranje membrana u embrionima viših životinja.
ili tako
FILEMBRIOGENEZA (od grčkog phylon - rod, pleme, embrion - embrion i geneza - porijeklo), evolucijska promjena u ontogenezi organa, tkiva i ćelija, povezana s progresivnim razvojem i redukcijom. Doktrinu filebriogeneze razvio je ruski evolucijski biolog A.N. Severtsov. Načini (metode) filebriogeneze razlikuju se po vremenu nastanka tokom razvoja ovih struktura.
Ako se razvoj određenog organa kod potomaka nastavi nakon faze na kojoj je završio kod predaka, dolazi do anabolije (od grčkog anabole - uzdizanje) - produžetka završne faze razvoja. Primjer je formiranje srca sa četiri komore kod sisara. Vodozemci imaju trokomorno srce: dva atrija i jednu komoru. Kod gmizavaca se u komori razvija septum (prva anabolija), ali kod većine je ovaj septum nekompletan – samo smanjuje miješanje arterijske i venske krvi. Kod krokodila i sisara razvoj septuma se nastavlja do potpunog odvajanja desne i lijeve komore (druga anabolija). Kod djece je ponekad, kao atavizam, interventrikularni septum nedovoljno razvijen, što dovodi do ozbiljne bolesti koja zahtijeva hiruršku intervenciju.
Produženje razvoja organa ne zahtijeva duboke promjene u prethodnim fazama njegove ontogeneze, stoga je anabolizam najčešći metod filebriogeneze. Faze razvoja organa koje prethode anabolizmu ostaju uporedive sa fazama filogenije predaka (tj. rekapitulacije) i mogu poslužiti za njegovu rekonstrukciju (vidi Biogenetski zakon). Ako razvoj organa u srednjim fazama odstupi od puta kojim se odvijala njegova ontogeneza kod njegovih predaka, dolazi do devijacije (od kasnolat. deviatio - odstupanje). Na primjer, kod riba i gmizavaca ljuske se pojavljuju kao zadebljanja epiderme i donjeg sloja vezivnog tkiva kože - korijuma. Postepeno se zgušnjavajući, ovaj anlage se savija prema van. Zatim kod riba korij okoštava, formirane koštane ljuske probijaju epidermu i pomiču se na površinu tijela. Kod gmizavaca, naprotiv, kost se ne formira, ali epiderma postaje keratinizirana, formirajući rožnate ljuske guštera i zmija. Kod krokodila, korij može okoštati, formirajući koštanu osnovu rožnatih ljuski. Odstupanja dovode do dubljeg restrukturiranja ontogeneze od anabolizma, pa su rjeđa.
Najrjeđe se javljaju promjene u primarnim rudimentima organa - archallaxis (od grčkog arche - početak i allaxis - promjena). U slučaju odstupanja, rekapitulacija se može pratiti od nastanka organa do trenutka razvojne devijacije. Kod arhalaksije nema rekapitulacije. Primjer je razvoj tijela kralježaka kod vodozemaca. Kod fosilnih vodozemaca - stegocefalija i kod modernih vodozemaca bez repa, tijela pršljenova su formirana oko tetive od nekoliko, obično po tri na svakoj strani tijela, odvojenih lanaca, koji se zatim spajaju u tijelo pršljena. Kod repatih vodozemaca ovi anlage se ne pojavljuju. Osifikacija raste iznad i ispod, pokrivajući notohordu, tako da se odmah formira koštana cijev, koja, zadebljajući, postaje tijelo pršljena. Ova arhalaksija je razlog za pitanje porijekla repatih vodozemaca o kojem se još uvijek raspravlja. Neki znanstvenici vjeruju da potječu direktno od riba s režnjevima, bez obzira na druge kopnene kralježnjake. Drugi kažu da su se repasti vodozemci vrlo rano odvojili od drugih vodozemaca. Drugi pak, zanemarujući razvoj pršljenova, dokazuju blisku vezu repatih i bezrepih vodozemaca.
Smanjenje organa koji su izgubili adaptivni značaj nastaje i kroz filebriogenezu, uglavnom negativnim anabolizmom – gubitkom završnih faza razvoja. U ovom slučaju, organ se ili nedovoljno razvija i postaje rudiment, ili se podvrgava obrnutom razvoju i potpuno nestaje. Primjer rudimenta je ljudsko slijepo crijevo - nerazvijeni cekum; primjer potpunog nestanka je rep punoglavaca žabe. Tokom svog života u vodi, rep raste, na njegovom kraju se dodaju novi pršljenovi i mišićni segmenti. Tokom metamorfoze, kada se punoglavac pretvori u žabu, rep se rastvara, a proces se odvija obrnutim redoslijedom - od kraja do baze. Filembriogeneza je glavna metoda adaptivnih promjena u strukturi organizama tokom filogeneze.
Ulaznica 97
Principi (metode) filogenetskih transformacija organa i funkcija. Podudarnost strukture i funkcije u živim sistemima. Multifunkcionalnost Kvantitativne i kvalitativne promjene u funkcijama bioloških struktura.
.
Principi filogenetskih transformacija
Autoritet nazivaju istorijski razvijeni specijalizovani sistem tkiva, koji karakteriše razgraničenje, konstantnost oblika, lokalizacija, unutrašnja struktura cirkulatornih i inervacionih puteva, razvoj u ontogenezi i specifične funkcije. Struktura organa je često veoma složena. Većina njih je multifunkcionalna, tj. obavlja nekoliko funkcija istovremeno. Istovremeno, različiti organi mogu učestvovati u realizaciji bilo koje složene funkcije.Grupa organa sličnog porijekla koji se ujedinjuju radi obavljanja složene funkcije naziva se sistem(cirkulatorni, ekskretorni i dr.) Ako istu funkciju obavlja grupa organa različitog porijekla, naziva se aparata. Primjer je respiratorni aparat koji se sastoji od samih respiratornih organa i elemenata skeleta i mišićnog sistema koji obezbjeđuju respiratorne pokrete.
U procesu ontogeneze dolazi do razvoja i često zamjene nekih organa drugim. Organi zrelog organizma nazivaju se definitivan; organi koji se razvijaju i funkcionišu samo u embrionalnom ili larvalnom razvoju - provizorno. Primjeri privremenih organa su škrge larvi vodozemaca, primarni bubreg i embrionalne membrane viših kralježnjaka (amniote).
U istorijskom razvoju, transformacije organa mogu imati progresivan ili regresivan karakter. U prvom slučaju organi se povećavaju u veličini i postaju složeniji u svojoj strukturi, u drugom se smanjuju i njihova struktura postaje jednostavnija.
Ako dva organizma koja se nalaze na različitim nivoima organizacije imaju organe koji su izgrađeni prema jednom planu, smješteni na istom mjestu i razvijaju se na sličan način iz istih embrionalnih rudimenata, onda to ukazuje na odnos ovih organizama. Takvi organi se nazivaju homologno. Homologni organi često obavljaju istu funkciju (na primjer, srce riba, vodozemaca, gmizavaca i sisara), ali se u procesu evolucije funkcije mogu mijenjati (na primjer, prednji udovi riba i vodozemaca, gmazova i ptica).
Kada nepovezani organizmi žive u istom okruženju, oni mogu razviti slične adaptacije, koje se manifestiraju u izgledu slično organi. Slični organi obavljaju iste funkcije, ali se njihova struktura, lokacija i razvoj oštro razlikuju. Primjeri takvih organa su krila insekata i ptica, udovi i vilični aparat artropoda i kralježnjaka.
Struktura organa striktno odgovara funkcijama koje obavljaju. Štoviše, u povijesnim transformacijama organa, promjena funkcija svakako je praćena promjenom morfoloških karakteristika organa.
Osnovni princip evolucije organskih struktura je princip diferencijaciju. Diferencijacija je podjela homogene strukture na zasebne dijelove, koji zbog različitih položaja, veza s drugim organima i različitih funkcija dobijaju specifičnu strukturu. Dakle, komplikacija strukture je uvijek povezana s usložnjavanjem funkcija i specijalizacijom pojedinih dijelova. Diferencirana struktura obavlja nekoliko funkcija, a njena struktura je složena.
Primjer filogenetske diferencijacije može biti evolucija cirkulacijskog sistema u tipu hordata. Tako je kod predstavnika podtipa bez lubanje strukturiran vrlo jednostavno: jedan krug cirkulacije krvi, odsustvo srca i kapilara u sistemu škržnih arterija.
Superklasa riba ima dvokomorno srce i škržne kapilare. Kod vodozemaca se po prvi put cirkulacijski sistem dijeli na dva kruga cirkulacije, a srce postaje trokomorno. Maksimalna diferencijacija je karakteristična za cirkulatorni sistem sisara, čije je srce četvorokomorno, a u sudovima se postiže potpuno razdvajanje venskog i arterijskog krvotoka.
Pojedini dijelovi diferencirajuće, prethodno homogene strukture, specijalizirani za obavljanje jedne funkcije, postaju funkcionalno sve više zavisni od drugih dijelova ove strukture i organizma u cjelini. Ova funkcionalna podređenost pojedinih komponenti sistema u cijelom organizmu naziva se integracija.
Četverokomorno srce sisara primjer je visoko integrirane strukture: svaki odjel obavlja samo svoju posebnu funkciju, koja nema smisla odvojeno od funkcija drugih odjela. Stoga je srce opremljeno autonomnim sistemom funkcionalne regulacije u obliku parasimpatičkog atrioventrikularnog nervnog čvora i istovremeno je strogo podređeno neurohumoralnom regulacijskom sistemu tijela u cjelini.
Tako se istovremeno sa diferencijacijom uočava i podređenost delova integralnom sistemu tela, tj. proces integracije.
U procesu evolucije, prirodno je da emergence nove strukture i njihove nestanak. Zasnovan je na principu diferencijacije, koji se manifestira na pozadini primarne polifunkcionalnosti i sposobnosti funkcija da se kvantitativno mijenjaju. Svaka struktura u ovom slučaju nastaje na osnovu prethodnih struktura, bez obzira na to na kom nivou organizacije živih bića se odvija proces filogeneze. Dakle, poznato je da se prije otprilike 1 milijardu godina originalni globinski protein, nakon umnožavanja originalnog gena, diferencirao u mio- i hemoglobin - proteine koji su dio mišićnih, odnosno krvnih stanica, i stoga diferencirani po funkciji. U filogenezi centralnog nervnog sistema hordata može se uočiti i diferencijacija i promena funkcija struktura: mozak se formira od prednjeg kraja neuralne cevi. Na isti način se formiraju nove biološke vrste u obliku izolovanih populacija izvornih vrsta (vidi § 11.6), a nove biogeocenoze - zbog diferencijacije već postojećih (vidi § 16.2).
S obzirom na to da će se u nastavku razmatrati filogenija određenih organskih sistema, detaljnije ćemo se zadržati na obrascima pojave i nestajanja organa. Primjer emergence organa je porijeklo materice placentnih sisara iz parnih jajovoda. Odugovlačenjem embrionalnog razvoja sisara javlja se potreba za dužim zadržavanjem embriona u majčinom tijelu. To se može izvesti samo u kaudalnim dijelovima jajovoda, čija je šupljina proširena, a zid je diferenciran na način da je posteljica pričvršćena za njega, osiguravajući odnos između majke i fetusa. U procesu prirodne selekcije očuvali su se i uspješno razmnožavali prije svega oni sisari u čijim se tijelima najduže razvilo potomstvo. Kao rezultat toga, nastao je novi organ - materica, koja embrionu obezbeđuje optimalne uslove za intrauterini razvoj (videti odeljak 14.5.3) i povećava stopu preživljavanja odgovarajuće vrste.
U nastanku tako složenijeg i specijaliziranijeg organa kao što je oko, primjećuju se isti obrasci. Formiranje organa vida, kao i svih čulnih organa, zasniva se na epitelnim ćelijama kože, među kojima se razlikuju receptorske ćelije, posebno fotosenzitivne ćelije. Njihovo spajanje u grupe dovodi do pojave primitivnih odvojenih organa vida, omogućavajući životinjama samo procjenu osvjetljenja. Uranjanje ovakvog organa osjetljivog na svjetlost pod kožu osigurava sigurnost osjetljivih stanica, ali se vizualna funkcija može postići samo zbog pojave transparentnosti integumenta. Osetljivost na svetlost primitivnog organa vida povećava se sa zadebljanjem prozirnih omotača i njihovim sticanjem sposobnosti da prelamaju svetlost i fokusiraju njene zrake na osetljive ćelije oka. Složen organ zahtijeva pomoćni aparat - zaštitne strukture, mišiće koji ga pokreću itd. Povećani nivo složenosti organizacije oka nužno je praćen i komplikacijom regulacije njegovih funkcija, što se izražava u jačanju njegove integracije kao integralnog sistema.
Nestanak ili redukcija organa u filogeniji može biti povezana s tri različita razloga i ima različite mehanizme. Prvo, organ koji je ranije obavljao važne funkcije može se pokazati štetnim u novim uvjetima. Prirodna selekcija djeluje protiv toga, a organ vrlo brzo može potpuno nestati. Malo je primjera takvog direktnog nestanka organa. Stoga su mnogi insekti malih oceanskih ostrva bez krila zbog stalnog uklanjanja letećih jedinki iz njihove populacije vjetrom. Nestanak organa češće se opaža zbog njihove zamjene novim strukturama koje obavljaju iste funkcije s većim intenzitetom. Tako, na primjer, kod gmizavaca i sisara, prednji pupoljci i primarni bubrezi nestaju, a funkcionalno ih zamjenjuju sekundarni bubrezi. Na isti način, kod riba i vodozemaca, notohorda je pomjerena kičmom.
Najčešći put do nestanka organa je kroz postepeno slabljenje njihovih funkcija. Takve situacije obično nastaju kada se životni uslovi promijene. Organ koji ne obavlja gotovo nikakve funkcije izmiče kontroli prirodne selekcije i obično pokazuje povećanu varijabilnost. Nastale promjene uzrokuju prekid korelativnih veza sa drugim dijelovima tijela. Zbog toga takav organ često postaje štetan i prirodna selekcija počinje djelovati protiv njega.
U medicinskoj praksi je opšte poznato da se rudimentarni organi kod ljudi odlikuju velikom varijabilnosti. Treći veliki kutnjaci, ili „umnjaci“, na primjer, karakteriziraju ne samo značajna varijabilnost u strukturi i veličini, već i različiti periodi nicanja, kao i posebna osjetljivost na karijes. Ponekad uopće ne eruptiraju, a često se nakon izbijanja potpuno unište u narednih nekoliko godina. Isto važi i za vermiformni dodatak cekuma (apendiks), koji normalno može imati dužinu od 2 do 20 cm i biti različito lociran (iza peritoneuma, na dugom mezenteriju, iza cekuma itd.). Osim toga, upala slijepog crijeva (apendicitis) javlja se mnogo češće od upalnih procesa u drugim dijelovima crijeva.
Proces redukcije organa je suprotan njegovoj normalnoj morfogenezi. Prije svega, ispadaju oznake takvih dijelova organa, koji se obično zadnji formiraju. Kada su udovi osobe nedovoljno razvijeni, najčešće su prve nedovoljno razvijene falange 1. i 5. prsta, koje se posljednje razvijaju. Kod kitova, koji su zbog slabljenja funkcija u filogenezi potpuno lišeni stražnjih udova, još uvijek su ostali rudimenti elemenata karličnog pojasa koji se formiraju vrlo rano u procesu morfogeneze.
Istraživanja genetske osnove redukcije organa pokazala su da strukturni geni koji reguliraju morfogenezu ne nestaju, dok geni koji reguliraju vrijeme formiranja rudimentarnih organa, odnosno geni odgovorni za fenomen induktivnih interakcija u embrionu u razvoju, prolaze kroz značajne promjene. Zaista, kada se mezodermalni materijal dna usne šupljine embriona guštera presađuje u usnu šupljinu pilića u razvoju, moguće je da potonji formira zube tipične strukture, a transplantacija mezoderma kože gušter ispod epiderme leđa pilića dovodi do stvaranja tipičnih rožnatih ljuski u njemu umjesto perja.
Sličnosti između embrija predstavnika različitih grupa kralježnjaka
Razvoj svih životinja počinje od jedne ćelije.
Organogeneza se odvija u određenom nizu. Kod hordata počinje With formiranje rudimenta notohorde i nervnog sistema. Na dorzalnoj strani embrija grupa ćelija ektoderma se odvaja u dugačku ploču. Ove stanice počinju se aktivno dijeliti, uranjajući u tijelo embrija i formirajući žlijeb, čiji se rubovi postupno približavaju, a zatim se zatvaraju, formirajući primarnu neuralnu cijev.
Osim nervnog sistema, iz ektoderma nastaju i kožne žlijezde, zubna caklina, kosa, nokti i epitel kože. . Endoderm stvara tkiva koja oblažu crijeva i respiratorni trakt , formira jetru i gušteraču. Od mezoderma se formiraju mišići, hrskavica i koštani skelet, organi za izlučivanje , reproduktivni i cirkulatorni sistem tijela .
Tokom procesa embriogeneze, postoji bliska interakcija između dijelova embrija u razvoju: rudiment jednog organa ili sistema organa određuje (inducira) lokaciju i vrijeme formiranja drugog organa ili sistema organa.
Diferencijacija embrionalnih ćelija ne nastaje odmah, već u određenoj fazi razvoja. U ranim fazama cijepanja, ćelije embrija još nisu specijalizirane, pa svaka od njih može dati povod za cijeli organizam. Ako se iz bilo kojeg razloga ove ćelije razdvoje, formiraju se dvije identične ćelije. embrion, koji sadrže identične genetske informacije, od kojih se svaka razvija u punopravnu jedinku. Kao rezultat, rađaju se identični ili monozigotni blizanci. U ljudskoj populaciji, to su jedini ljudi koji imaju identičan genotip i kopije su jedni drugih,
Međusobni utjecaj dijelova embrija dokazan je u brojnim eksperimentima. njemački istraživači Hans Spemann i Hild Mangold uzeo je od embriona trimota u ranoj fazi gastrule dio dorzalne strane tijela, iz kojeg V dalje, trebalo je razviti notohord i mezoderm, te je presađen na ventralnu stranu druge gastrule . IN Kao rezultat toga, na trbušnoj strani drugog embrija, formirana je dodatna neuralna cijev od ćelija koje su trebale da izazovu kožu. Ovaj fenomen se zove Embrionalna indukcija.
Stvaranje komparativne embriologije
K.M. Bare je prvi primenio embriološki kriterijum u sistematici i potkrepio njegov značaj. Upoređujući metode razvoja životinja, naučnik je došao do zaključka da postoje 4 tipa organizacije - tip kralježnjaka, izduženi ili zglobni tip, masivni tip i periferni ili zračeći tip. Svaki tip odgovara posebnom planu razvoja. Unutar svakog tipa mogu se razlikovati različiti nivoi razvoja, koji odgovaraju klasama. Životinje se međusobno razlikuju i po tipu ontogeneze i po stepenu heterogenosti.
Tako su se sredinom 19. stoljeća, s jedne strane, ocrtavale razlike u ontogenezi životinja različitih tipova i klasa. S druge strane, postalo je jasno da embrionalni razvoj vrlo različitih organizama teče formiranjem zametnih slojeva, što nam omogućava da govorimo o homolognim strukturama kod predstavnika različitih grupa. Postalo je moguće uporediti podatke iz embriologije i komparativne anatomije.
Početkom 19. vijeka. I. Meckel, L. Oken, M. Rathke, Gidemman iznijeli su tvrdnju da embrioni viših životinja prolaze u svom razvoju kroz sve faze koje odgovaraju odraslom stanju nižih životinja.
K.M. Bare, pažljivo proučavajući individualni razvoj predstavnika različitih grupa kralježnjaka i beskičmenjaka, došao je do zaključka da se mogu porediti samo životinjski embriji. U procesu embrionalnog razvoja prvo se pojavljuju karakteri tipa, zatim klasa, red, porodica, rod i vrsta. Što su embriji mlađi koji pripadaju različitim klasama istog tipa, to su sličniji jedni drugima. Drugim riječima, predstavnici različitih grupa, na primjer, klasa podtipa kralježnjaka, u ranim fazama ontogeneze obično su sličniji od odraslih jedinki. Ovaj obrazac se zove Zakon germinativne sličnosti .
Rezultati embrioloških studija K.M. Korišteni su Baer C. Darwin kao jedan od dokaza evolucije.
Komparativna embriologija
Von Baer (1792-1867), proučavajući embrionalni razvoj kod predstavnika različitih grupa kralježnjaka, otkrio je iznenađujuće strukturne sličnosti u svim ovim grupama, posebno u fazama cijepanja, gastrulacije i diferencijacije organizma u nastajanju.
Haeckel (1834-1919) je sugerirao da ova sličnost ima evolucijski značaj. Formulirao je zakon rekapitulacije, prema kojem „ Ontogeneza ponavlja filogenezu", tj. faze kroz koje organizam prolazi u procesu svog razvoja ponavljaju evolucijsku istoriju grupe kojoj pripada. Samo proučavanje ranih embriona bilo kojeg kralježnjaka pokazuje da je nemoguće odrediti grupu kojoj pripadaju. Tek u relativno kasnim fazama razvoja embrij počinje imati neku sličnost s odgovarajućim odraslim oblikom. Početna sličnost između embriona objašnjava se činjenicom da su svi oni, a samim tim i klase kojima pripadaju, imali zajedničkog pretka. Zakon rekapitulacije, međutim, ne može se prihvatiti bezuslovno, budući da nijedan živi organizam ne može pokazati sve znakove svojih pretpostavljenih evolucijskih predaka. Ali čini se vjerojatnim da organizmi zadržavaju razvojne mehanizme naslijeđene od svojih predaka. Stoga je moguće da će u različitim fazama razvoja dati organizam imati osobine strukturne sličnosti sa embrionima oblika predaka. Naknadne adaptacije na druge uslove sredine i stilove života menjaju dalji tok razvoja. Kao što pokazuju zapažanja, što su bliže grupe kojima su dva data organizma dodijeljena na osnovu zajedničkih homoloških struktura, to duže ostaje njihova sličnost u embrionalnim fazama. Organizmi prilagođeni određenom načinu života i određenom staništu, koji nisu tipični za veliku grupu kojoj pripadaju, u procesu embrionalnog razvoja manje su slični ostalim pripadnicima ove grupe.
BEROV ZAKON- grupa odredbi koje postuliraju sličnost ranih faza razvoja embrija predstavnika različitih klasa kralježnjaka. K. E. Baer (ruski embriolog koji je živeo u 19. veku) verovao je da prvo embrioni razvijaju karaktere tipa, zatim klasu, red i, konačno, u kasnijim fazama razvoja - individualne karaktere koji karakterišu specijalizaciju taksona, tj. embrioni različitih klasa su u početku slični, a zatim odstupaju jedan od drugog u svom razvoju.
MUELLER-HOCKEL ZAKON (biogenetski) - navodi da je ontogenija kratko ponavljanje filogenije. Zakon je formulisao E. Haeckel na osnovu ideja Charlesa Darwina i istraživanja F. Mullera. Početkom 20. vijeka. zakon je dopunjen stavom A. N. Severtsova da se u ontogenezi organizama ne ponavljaju čitavi filogenetski stadijumi, već je moguća samo rekapitulacija (kratko i sažeto ponavljanje) pojedinačnih karakteristika i procesa.
Zakon germinativne sličnosti
Istraživači ranog 19. veka. po prvi put su počeli obraćati pažnju na sličnost faza razvoja embriona viših životinja sa fazama složenosti organizacije koje vode od nisko organiziranih oblika ka progresivnim. Upoređujući faze razvoja embriona različitih vrsta i klasa hordata, K. Baer je došao do sljedećih zaključaka.
1. Embrioni životinja istog tipa su slični u ranim fazama razvoja.
2. Dosljedno se kreću u svom razvoju od opštijih karakteristika tipa ka sve specifičnijim. Konačno, razvijaju se znakovi koji ukazuju da embrij pripada određenom rodu, vrsti i, konačno, individualnim osobinama.
3. Embrioni različitih predstavnika istog tipa postepeno se odvajaju jedan od drugog
Sličnosti embriona i embrionalna divergencija karaktera (K. Baerov zakon)
biogenetski zakon (E. Haeckel i F. Müller) A. N. Severtseva
1. Uvod
4. Radovi Severcova A.N.
Bibliografija
1. Uvod
Prirodni filozofi 18. i 19. vijeka. Rekli su da je život u osnovi jedan. Ovo jedinstvo je izraženo u njihovom paralelizmu između embrionalnog razvoja pojedinca i lestvice živih bića, koja datira još od Aristotela. Prema konceptu lestvice živih bića, svi objekti koji postoje u prirodi su karike u neprekidnom lancu koji povezuje neorganske tvorevine sa nizom živih oblika sve složenije. Iz nežive prirode dolazi do postupnog prijelaza na biljke, a zatim na tako jednostavne životinje kao što su spužve, zatim na insekte, ribe, ptice, sisavce i na kraju na ljude. Ovaj obrazac je bio statičan i trebalo bi ga tumačiti kao evolucijski. Jednostavno je predstavljao plan prema kojem je Bog stvorio svijet.
Prema zakonu paralelizma, poznatom kao Meckel-Serresov zakon (po imenima njegova dva tvorca: Meckel u Njemačkoj, Serres u Francuskoj), svako živo biće u svom embrionalnom razvoju ponavlja odrasle oblike životinja koje stoje na nižim stepenicama merdevine živih bića. Nasuprot tome, niže životinje predstavljaju trajne faze ličnosti evolucijski naprednijih oblika. Zakon paralelizma, kao i ljestve živih bića, nije sadržavao ništa evolutivno. Detaljna analiza takvog paralelizma omogućila je Karlu Baeru da formuliše zakon germinalne sličnosti.
2. Sličnosti embriona i embrionalna divergencija karaktera (K. Baerov zakon)
Karl Maksimovič Baer (aka Karl Ernst von Baer) rođen je 1792. godine u Estoniji, a ovdje, u Dorpatu (danas Tartu), završio je svoj životni put 1876. godine. Konigsberški period Baerove aktivnosti povezan je s embriologijom. Radeći na Univerzitetu u Königsbergu, svoja istraživanja, koja su u velikoj mjeri postavila temelje embriologije kao nauke, vodio je u atmosferi dominacije prirodne filozofije.
On je, slijedeći Wolfa i Pandera, proučavao embrionalni razvoj pilića, a zatim i niz drugih predstavnika različitih klasa kralježnjaka. Otkrio je jaje kod sisara, proučavao razvoj fetalnih membrana i klica; otkrio je i opisao blastulu, fazu embrionalnog razvoja kod životinja. Sva ova otkrića napravio je u dobi od 25-36 godina. Baer je provodio dane i noći, sedmice i mjesece u laboratoriji.
Šta je mladog istraživača toliko fasciniralo? Zašto se on, po njegovim riječima, “pretvorio u neku vrstu rakova pustinjaka, nikad ne napuštajući oklop koji je jednom izabrao”? Baer je bio u mogućnosti da iznova pogleda na dugo poznati fenomen sličnosti u embrionalnim stadijima životinja koje pripadaju različitim klasama kralježnjaka.
Baer je bio prvi koji je identificirao tri oblika diferencijacije u embriologiji: cijepanje na zametne slojeve, ili „primarno odvajanje“; nastanak tkiva, ili „čisto-logička izolacija“ (ovo je napisano deceniju i po pre stvaranja ćelijske teorije!); nastanak organa ili „morfološko odvajanje“. Na osnovu rezultata svog rada na komparativnoj embriologiji napravio je niz generalizacija koje su pokazale potpunu besmislenost ideje da životinje u svom razvoju ponavljaju sve stepenice ljestvice živih bića.
Baer je, poput Cuviera, koji je 1812. godine stvorio sistem klasifikacije koji je precrtao ljestvicu živih bića, primijetio da ne postoji jedna sekvencijalna serija, već četiri osnovna plana za strukturu životinja. Ova četiri plana se jasno odražavaju u njihovom razvoju. Na primjer, notohorda* i neuralna cijev karakteristični za kičmenjake nastaju u ranim fazama razvoja i stoga je „embrion kičmenjaka od samog početka kralježnjak, i nijedan period ne odgovara beskičmenjaku. Embrioni kralježnjaka su slični samo drugim embrionima kralježnjaka; Baer poriče njihovu sličnost s odraslim osobama bilo koje druge životinje: "Zameci kralježnjaka* ne prolaze kroz trajne oblike nijedne (poznate) životinje u procesu svog razvoja." Činjenice, a ne predrasude, dovele su ga do evolucionih pogleda.
Obratite pažnju na to kako moderno zvuči ideja naučnika da „što su životni oblici više različiti jedan od drugog, to se mora zaroniti dalje u dubine istorije razvoja da bi se pronašle sličnosti među njima“. Međutim, pod riječima “istorija razvoja” Baer je mislio na historiju individualnog, odnosno embrionalnog, a ne historijskog razvoja.
Sumirajući uporedne podatke, Baer je 1828. objavio sljedeće osnovne generalizacije – svoje čuvene zakone, dijametralno suprotne teoriji paralelizma:
1. Zajedničko svakoj većoj grupi životinja formira se u embriju ranije od posebne...
2. Od opštijeg... nastaje manje opšte, i tako dalje, dok, konačno, ne nastane najposebnije...
3. Embrion svakog životinjskog oblika uopšte ne ponavlja druge životinjske oblike tokom razvoja, već se, naprotiv, odvaja od njih.
4. U osnovi, dakle, embrion najvišeg oblika životinje nikada nije sličan drugom obliku životinje, već samo njegovom embriju.
Nije teško uočiti polemičke prizvuke Baerovih zaključaka, posebno posljednja dva, usmjerena protiv teorije paralelizma. Međutim, one su potkrijepljene činjenicama, a dvije glavne odredbe, nazvane princip embrionalne sličnosti i princip specijalizacije individualnog razvoja, zadržale su svoj značaj do danas. Zaista, embrioni različitih vrsta su vrlo slični jedni drugima, a kako se razvijaju, ta sličnost se gubi. U ranim fazama razvoja pilećeg embriona - Baerovog omiljenog predmeta istraživanja - može se vidjeti samo da on pripada kralježnjacima, jer rani embrioni kralježnjaka svih klasa izgledaju gotovo isto; malo kasnije može se identificirati kao ptica, a tek kasnije postaje očito da je to buduća kokoška.
Puteve i obrasce individualnog razvoja, zakone ontogeneze, pitanja odnosa ontogeneze i filogeneze potom su razvili Darwin, Müller i Haeckel, A. N. Severtsov. Ali svi su oni u svojim konstrukcijama polazili od činjeničnog temelja koji su u embriologiji stvorili radovi Karla Baera.
Ovaj izuzetni naučnik je značajna ličnost prirodne nauke 19. veka. Embriologija, ihtiologija, teriologija, komparativna anatomija, geografija, etnografija, lov i ribolov, istorija nauke, bibliografija - ovo nije potpuna lista onih oblasti znanja u kojima je Baer ostavio svoje tragove. Baer nije bio evolucionista, ali je svojim zakonom pripremio teren za evolucijsko tumačenje procesa ontogeneze. Već je Charles Darwin naglasio da proučavanje individualnog razvoja organizama baca svjetlo na njihovo porijeklo, budući da se „u embrionu može vidjeti nejasan portret pretka“. Ova pozicija je bila osnova poznatog biogenetskog zakona koji su ustanovili Ernst Haeckel i Fritz Müller.
3. Biogenetski zakon (E. Haeckel i F. Muller)
Pojavom teorije Charlesa Darwina o poreklu vrsta (1859.), teorija paralelizma J. Meckela dobila je evolucijsko objašnjenje: ljestve stvorenja prestale su biti statične, postalo je jasno da odražava evolucijsku težnju ka poboljšanju organizacije, i ontogeneza - individualni razvoj pojedinca - je proces evolucije. Istovremeno, postalo je moguće objasniti embrionalnu sličnost srodstvom (zajedničko porijeklo) organizama, ali je princip specijalizacije individualnog razvoja ostao neobjašnjen, a K. Baer je do kraja bio protivnik i I. Meckela i Charlesa Darwina. njegovog života. Dakle, opće ideje o ontogenezi su se pokazale kontradiktornima: s jedne strane, ovaj proces odražava evoluciju organizacije, s druge, samo sebe.
Osim toga, Charles Darwin, da bi objasnio evoluciju ranih faza razvoja, razvio je princip nasljeđivanja u odgovarajućoj dobi, prema kojem se osobina, jednom kada se pojavi u jednoj fazi ontogeneze, ima tendenciju da se pojavi kod potomaka u istoj fazi. , što omogućava da se izborom promijeni bilo koji stupanj individualnog razvoja . Ovaj obrazac nije objasnio ni paralelizam ni specijalizaciju razvoja. Kao rezultat toga, javila se potreba da se pomire svi kontradiktorni koncepti, a proučavanje odnosa između individualnog i istorijskog razvoja počelo je gotovo iznova.
Darwinove ideje razvio je Fritz Müller, koji je učinio mnogo u potkrepljivanju biogenetskog zakona. F. Müller (1821-1897) - njemački zoolog, koji je zauvijek otišao u Brazil 1852. godine. Nikada nije sreo ni Darwina ni Hekela. Iz provincijskog brazilskog grada Desterra, Müller je poslao malu knjigu pod naslovom “Za Darwina” (1864) za objavljivanje u Lajpcigu. U njemu je, na osnovu proučavanja ontogeneze viših rakova, došao do dva važna zaključka:
1. Ontogeneza potomaka može se nastaviti dalje od faze na kojoj je završila kod predaka. Tada organizacija potomaka postaje viša od organizacije njihovih predaka.
2. Ontogeneza potomaka može, u srednjim fazama, skrenuti sa puta kojim je išla među svojim precima. Tada će organizacija potomaka biti drugačija, ali ne viša od one njihovih predaka.
U ovom radu nisu razmatrani ni mehanizmi individualnog razvoja niti evolucija ontogeneze kao procesa. Govorili smo samo o evoluciji odraslog stadija i razmatrali kako promjene u toku ontogeneze utiču na definitivnu strukturu. Razlozi promjena u toku ontogeneze nisu dotaknuti.
Pa ipak, ovaj rad F. Mullera bio je važan korak naprijed u razvoju teorije evolucije ontogeneze. Najprije je pokazao da evolucijski novi oblici nastaju promjenom toka individualnog razvoja karakterističnog za njihove pretke, odnosno promjene u ontogenezi su primarne u odnosu na filogenetske promjene. Drugo, pokazalo se da promjene u toku ontogeneze mogu nastati na različite načine. Treće, na osnovu Müllerovih podataka može se tvrditi da ako se ontogeneza potomaka nastavi dalje od definitivne faze predaka, iz ove, koja je već postala pretposljednja, može se suditi o organizaciji predaka, a time i o nastavku proučavanih oblika. Kao rezultat, djelomično je objašnjena teorija paralelizma i potkrijepljena uloga embriologije u rekonstrukciji filogenije. Treba samo napomenuti da je Muller razmatrao ontogenezu cijelog organizma, a ne njegovih dijelova (pojedinačne organizme i histogenezu*).
Ideje F. Mullera poslužile su kao osnova za dalji razvoj problema. Koristio ih je Ernst Haeckel (1834-1919), koji je odigrao ogromnu ulogu u propagandi i daljem razvoju evolucionog učenja. Haeckel i Müller razvili su ideju da su uzastopne promjene oblika jedinke tokom individualnog razvoja posljedica filogeneze, odnosno razvoja roda kojem pripada određena vrsta životinje.
Ontogeneza je kratko i brzo ponavljanje filogeneze, zbog nasljednosti i adaptacije. Ovaj obrazac je Haeckel (1866.) nazvao osnovnim biogenetičkim zakonom. Potpuna formulacija ovog zakona je sljedeća: razvoj embrija (individualni ili ontogenetski niz razvoja, ontogeneza) je komprimirano, skraćeno ponavljanje razvoja cijele grupe oblika koji su proizašli iz jednog zajedničkog korijena (tj. ponavljanje razvoja filetske, ili paleontološke, filogenije); što je ponavljanje potpunije, to je izvorni razvoj predaka (palingeneza) sačuvan zbog stalnog nasljeđivanja. Ponavljanje je manje potpuno, što je više kasnijih poremećaja (cenogeneza) uvedeno zahvaljujući različitim adaptacijama.
Prema ovom zakonu, embrioni viših životinja su slični odraslim nižim precima. Jaja svih životinja su "identična". Evolucija se dešava dodavanjem novih faza na kraju razvoja. Da se ništa ne meša, onda bi se evoluciona istorija vrste tačno ponovila u individualnom razvoju, što znači da bismo tokom sati, nedelja ili meseci razvoja embriona ove ili one životinje mogli gledati, slikovito rečeno, „film“ o svojoj istorijskoj prošlosti. Gledajući „film“ o individualnom razvoju sisara, vidjeli bismo faze koje odgovaraju odraslim oblicima beskičmenjaka, a zatim riba, vodozemaca i gmizavaca.
Međutim, ispoljavanje u nepromijenjenom obliku primitivnih, antičkih, palingenetskih karaktera (od grčkog palaios - drevni; palingenesis - karakteri koji su prešli u ontogenezu životinje iz njene filogeneze) otežava cenogeneza, različiti karakteri koji su nastali u ontogenezi kao adaptacija na uslove života embrija larvi i odraslih životinja. Primjeri koenogeneze uključuju amnion, horion, alantois Amniota i ogromne žlijezde koje se okreću larvi insekata.
Proces ponavljanja filogenetskih karaktera u ontogenetskom razvoju naziva se rekapitulacija. Tipičan primjer rekapitulacije daje akademik A. N. Severtsov u svojoj knjizi "Morfološki obrasci evolucije", objavljenoj 1913. u Njemačkoj. Znamo da su kod modernih vodozemaca bez repa, tibija i fibula spojene zajedno kao kod odraslih, ali kod punoglavaca su odvojene. Stegocefali, od kojih su evoluirali moderni vodozemci, također su imali dvije odvojene potkoljenice. Shodno tome, prisustvo odvojenih kostiju tibije kod punoglavaca može se smatrati rekapitulacijom jedne od karakteristika stražnjih udova karakterističnih za skelet predaka.
Rekapitulacija nije ograničena na morfološke karakteristike. Mogu se identifikovati i tokom ontogenetskog razvoja funkcija različitih organa i tkiva. Poznato je da su se tokom evolucije kralježnjaka postepeno gubili enzimi neophodni za razgradnju mokraćne kiseline. Tako je kod nekih gmizavaca i ptica krajnji produkt metabolizma dušika mokraćna kiselina, kod vodozemaca i većine riba urea, a kod primarnih beskičmenjaka amonijak. Pokazalo se da ptičji embrion luči amonijak u ranim fazama razvoja, ureu u kasnijim fazama, a mokraćnu kiselinu tek u zadnjim fazama. Slično, kod punoglavaca krajnji produkt metabolizma je amonijak, a kod žaba urea.
Biogenetski zakon postao je srž Hekelove teorije rekapitulacije. Prema ovoj teoriji, ontogeneza uključuje dvije vrste karaktera: palingenezu - karaktere filogenetskih dalekih predaka i koenogenezu - sekundarne karaktere koji su nastali kao rezultat prilagođavanja određenim uvjetima embrionalnog i larvalnog života. Tipična koenogeneza su, na primjer, trahealne škrge larvi vretenaca i majmuna: u vodenom načinu života one služe kao privremeni respiratorni organi.
Haeckel je pridavao primarnu ulogu palingenetičkim karakterima naslijeđenim od dalekih predaka u razjašnjavanju filogenetskih odnosa. Proučavanje cenogeneze u ovom aspektu je beskorisno i samo prikriva ispoljavanje palingenetičkih karakteristika. Sa ove tačke gledišta, cenogeneze su „neugodne distorzije“ palingeneze.
U Hekelovoj teoriji bilo je mnogo propusta, a glavni je bio da se istorija odraslog organizma posmatrala izolovano od istorije embriona. Uprkos tome, ne može se potcijeniti ogroman značaj biogenetskog zakona u razvoju evolucijskog učenja. S daljnjim istraživanjem, međutim, pokazalo se da je sve teže uklopiti nagomilane činjenice u prokrustovo ležište usko formulirane teorije. Kritike su se pojačavale, ponekad su dostizale tačku potpunog poricanja biogenetskog zakona i samog principa rekapitulacije. Činilo se da se iscrpio i da prelazi u polje istorije nauke sa njenim neizbežnim greškama.
4. Radovi A. N. Severtsova
Akademik Aleksej Nikolajevič Severcov (1866-1936) uspeo je da prevaziđe ove pogrešne stavove. Teorija filebriogeneze koju je predložio obnovila je biogenetski zakon i princip rekapitulacije, dajući im potpuno novo svjetlo. A. N. Severtsov je razvijao svoju teoriju čitavog života.
Treba napomenuti da su A. N. Severtsov i njegova škola svoju pažnju usmjerili na zakone makroevolucije. Najraniji opšti biološki govor Severcova - govor "Evolucija i embriologija" - datira iz 1910. godine; 1922. objavio je "Studije o teoriji evolucije. Individualni razvoj i evolucija". Ovi radovi daju kritiku biogenetskog zakona u njegovom hekelovskom mehanističkom obliku, na osnovu specifičnih istraživanja A. N. Severcova i njegove škole. Raspravljajući o vječnom problemu prilagodljivosti i neutralnosti likova, Severcov je predložio razliku između primarnih adaptivnih promjena (protalaksija) i sekundarnih promjena koje su nastale zbog korelativne varijabilnosti (deutalaksa). Severtsov je skrenuo pažnju na postojanje filogenetskih promjena povezanih s promjenama u toku embrionalnog razvoja - takve adaptacije je nazvao filebriogenezom.
Govoreći o odnosu onto- i filogeneze, Severtsov je napisao: „Biogenetski zakon Müller-Haeckel se ne može smatrati opovrgnutim - on sadrži vrlo značajnu količinu istine, iako ne i potpunu istinu: odnos između filogenije i ontogeneze je mnogo više. složeniji nego što se do sada mislilo, i nisu ograničeni na odnose koje je otkrio Müller i razvio Haeckel.” Severcov je ukazao na razlike u vremenu pojavljivanja novih likova u ontogenezi i ustanovio da novi likovi ne „napune“ uvek niz uzastopnih faza koje se javljaju u ontogenezi. U nekim slučajevima ovaj niz je sačuvan, dok se u drugim može transformisati čitav niz faza ontogeneze.
"Morfološki obrasci evolucije" glavni su teorijski rad A. N. Severtsova. Objavljena je u Rusiji 1939. godine, nakon smrti naučnika. Posjedujući ogromno iskustvo u komparativnim anatomskim i embriološkim istraživanjima, Severcov je u ovoj knjizi dao široku sliku makroevolucije, njenih obrazaca i metoda proučavanja.
U prvim poglavljima ispitao je zadatke i metode filogenetskog istraživanja i opisao teoriju monofiletske (jednokorijenske) evolucije kao zadovoljavajuće činjenične i teorijske konstrukcije. Uprkos očiglednoj pristrasnosti prema proučavanju evolucije viših taksona, Severcov je takođe obratio pažnju na prilagodljivost karaktera malih sistematskih grupa. Smatrao je mogućim postojanje selektivno indiferentnih karaktera, ali je vjerovao da se oni šire u populaciji zbog njihove korelacije sa selektivno bitnim karakterima. Šest godina prije objavljivanja knjige Teodosija Grigorijeviča Dobžanskog "Genetika i porijeklo vrsta", A. N. Severtsov je dao blisku definiciju vrste: ujedinjujemo slične životinje u jednu vrstu na osnovu biološke osobine, odnosno na osnovu njihove sposobnosti parenja jedni s drugima i stvaraju plodno potomstvo"
Promjene u individualnom razvoju organizama ili u razvoju pojedinih organa, koje se uporno prenose na potomstvo, nove karakteristične osobine mogu nastati na početku embrionalnog razvoja, na početku formiranja odgovarajućih organa - došao je do ovog zaključka Severtsov. Ova vrsta filebriogeneze naziva se arhalaksija. U drugim slučajevima, primetio je Severcov, promene mogu uticati na bilo koje srednje faze u razvoju organa (ili celog organizma). Ova vrsta promjene naziva se devijacija. Konačno, promjene mogu nastati u završnim fazama razvoja organizama ili u završnim fazama razvoja organa. Ove promjene se nazivaju anabolizam (tj. prijelaz u završne faze razvoja).
Dakle, prema Severcovu, različite karakteristike odrasle životinje se formulišu u različito vreme tokom njene ontogeneze i ontogeneze organa, a zatim nepromenjene prelaze u stanje odrasle osobe kao definitivne karakteristike; dodavanja se dešavaju preko definitivnog stanja karaktera, koje je, razvijajući se u ontogenezi, istovremeno i „embrionalno“ i „odraslo“.
Severcov je stvorio hipotezu o nastanku i evoluciji ontogeneze višećelijskih životinja, a ideje o anabolizmu i na njima zasnovane rekapitulacije su od velikog značaja u njoj. Kao pristalica hipoteza o nastanku višećelijskih organizama iz jednoćelijskih kolonija, Severcov sugerira da je kod predaka modernih višećelijskih organizama, odnosno onih koji su slični živim biljkama Pandorina morum ili Volvox, ontogeneza bila kratka i sastojala se od superponiranih faza tokom razvoj kolonija; iz jednoćelijske faze razvila se dvoćelijska (prva nadgradnja), iz dvoćelijske faze četveroćelijska faza (druga nadgradnja) itd.
U Volvoxu, kroz seriju takvih supersa, organizam se razvija do sferične kolonije, koja se može sastojati od više od 20.000 ćelija, a zatim dolazi do faze (super) čiste diferencijacije (određenog niza tipova ćelija). Kod životinja kao što su niži koelenterati, ontogeneza se produžila. Hidra, prema Severtsovu, ima faze u ontogenezi koje odgovaraju fazama ontogeneze kolonijalnih oblika - periodu fragmentacije. Ovim fazama se dodaju i faze formiranja dvoslojnog embrija - gastrule, zatim formiranje polipa sa ektodermom, želučanom šupljinom i pipcima. Ovakvim fazama „superstrukture“ dodaje se samo faza rasta i čista diferencijacija (formiranje ubodnih i drugih ćelija). Tako je ontogeneza hidroida proširena za nekoliko novih faza dodatih konačnom stanju kolonijalnih oblika jednoćelijskih organizama.
Evolucijske promjene u ontogenezi koje dovode do pojave novih likova su pozitivna filebriogeneza. Postoji i negativna filebriogeneza, koja rezultira potpunom ili djelomičnom redukcijom organa ili njegovih dijelova. A. N. Severtsov je razlikovao dvije vrste redukcija: rudimentaciju i afaneziju.
Kod rudimentacije dolazi do pojednostavljenja organa od samih početnih faza njegovog embrionalnog razvoja, ili, kao rezultat retardacije, dolazi do kasnijeg formiranja organa i, kao rezultat, do njegove nerazvijenosti. U potonjem slučaju nestaju završni stadijumi, a kod odraslih oblika ovaj organ poprima juvenilni izgled. Primjer takve rudimentacije su modificirani udovi zmijolikih guštera: u Seps halcidu formiraju se samo tri prsta i udovi su embrionalne prirode.
S drugom vrstom redukcije - afanizijom - organ razvijen kod predaka u potpunosti nestaje kod njihovih potomaka, ali je embrionalni anlage ipak očuvan. Primjer afanizije je atrofija repa kod punoglavaca, koji nestaje tijekom metamorfoze kao rezultat resorpcije, za razliku od predaka bezrepih vodozemaca, kod kojih je rep bio razvijen iu odraslim oblicima. Na isti način izvrsno razvijeni zubi u mlađi modernih jesetri potpuno nestaju kod odraslih jedinki zbog promjene prehrane, međutim, kod predaka Acipenseridae zubi su se razvili i kod odraslih riba.
Dakle, prema teoriji A. N. Severtsova, različite promjene u ontogenezi dovode ne samo do pojave novih likova u odraslim oblicima, već i do njihovog nestanka na temelju pozitivne ili negativne filebriogeneze.
Raznolikost adaptivnih promjena u organima, koje se formiraju na različite načine, pažljivo je analizirao A. N. Severtsov. U svojoj klasifikaciji načina (metoda) filogenetskih promjena u organima polazio je od funkcionalnog principa, smatrajući morfološke promjene organa „načinom razvoja aktivne funkcije ili pasivne adaptacije u smjeru korisnom za cijeli organizam“. Dakle, kada je govorio o promjenama u bilo kojem dijelu tijela, Severtsov je uzeo u obzir integritet cijelog organizma i njegovu povezanost sa uvjetima okoline.
Teorija filebriogeneze sasvim zadovoljavajuće objašnjava evoluciju strukture organizama na svim nivoima - od ćelije do pojedinca iu svim fazama ontogeneze - od zigota* do odrasle životinje. Svaka ontogeneza i bilo koji njen stadijum može se smatrati složenim sistemom, prepletom filebriogeneze organa, tkiva i ćelija. Evolucija individualnog razvoja odvija se kroz evoluciju određenih organskih, čistih i citogeneza.
A. N. Severtsov je imao veliku školu, njegov uticaj se osećao ne samo u našoj zemlji. Ne samo F. G. Dobzhansky i N. V. Timofeev-Resovsky, već i B. Rensch i A. Remane bili su upoznati s njegovim idejama. U svojim kasnijim radovima, A. N. Severtsov se okrenuo genetskim podacima i podržao borbu genetičara protiv neolamarkizma. Međutim, Severcovljeve radove ignorirali su tako aktivni tvorci sintetičke teorije evolucije kao što su J. Huxley i J. G. Simpson. Indirektni uticaj Severcovljevih ideja na zapadne evolucioniste je van sumnje. Među njegovim brojnim učenicima bio je jedan koji je otišao mnogo dalje u biološkim naukama od svog učitelja. Ovo je I. I. Shmalgauzen, koji se cijeli svoj život, kao učitelj, bavio komparativnom anatomijom, embriologijom i općom filogenetikom. Postao je nasljednik rada A.N. Severtsova i jedan od prvih istraživača koji je izvršio sintezu klasičnog darvinizma, sa svim njegovim komponentama, sa genetikom.
Bibliografija
1. Veliki enciklopedijski rječnik. Velika ruska enciklopedija, Sankt Peterburg, Norint, 2001.
2. Vorontsov N. N. Razvoj evolucionih ideja u biologiji. M.: Izdavačka kuća. Odsjek UC DO MSU, Progres-Tradicija, ABF, 1999.
3. Reff R., Kofman T. Embrioni, geni, evolucija, M.: Mir, 1986.
4. Tokin B. P. Opća embriologija. M.: Viša škola, 1987.
5. Toshchenko V.P. Uvod u teoriju evolucije: kurs predavanja /pod. Ed. Polyansko-
6. Oxford Illustrated Encyclopedia, tom 2. Prirodni svijet. M., Izdavačka kuća "Infra-M", "Cijeli svijet", 1999.
7. A. S. Severtsov. Osnovne teorije evolucije. M., Izdavačka kuća Moskovskog državnog univerziteta, 1987
2.1 K. Baerov zakon sličnosti zametne linije
Istraživači ranog 19. veka. po prvi put su počeli obraćati pažnju na sličnost faza razvoja embriona viših životinja sa fazama složenosti organizacije koje vode od nisko organiziranih oblika ka progresivnim. Karl fon Baer je 1828. godine formulisao obrazac nazvan Baerov zakon: „Što se uporede raniji stadijumi individualnog razvoja, to se više sličnosti može naći.” Upoređujući faze razvoja embriona različitih vrsta i klasa hordata, K. Baer je došao do sljedećih zaključaka.
1. Embrioni životinja istog tipa su slični u ranim fazama razvoja.
2. Dosljedno se kreću u svom razvoju od opštijih karakteristika tipa ka sve specifičnijim. Konačno, razvijaju se znakovi koji ukazuju da embrij pripada određenom rodu, vrsti i, konačno, individualnim osobinama.
3. Embrioni različitih predstavnika istog tipa postepeno se odvajaju jedan od drugog (slika 1).
Slika 1. Sličnost embriona različitih klasa kičmenjaka u različitim fazama
K. Baer, budući da nije evolucionista, nije mogao povezati obrasce individualnog razvoja koje je otkrio s procesom filogeneze. Stoga su generalizacije koje je napravio imale značenje samo empirijskih pravila.
Naknadni razvoj evolucijske ideje omogućio je da se sličnost ranih embriona objasni njihovim istorijskim srodstvom, i njihovo sticanje sve više posebnih osobina uz postepenu izolaciju jedna od druge - stvarna izolacija odgovarajućih klasa, redova, porodica, rodova i vrsta u procesu evolucije.
Teorija evolucije koju je razvio Charles Darwin jasno je istakla fundamentalnu važnost problema ontogenetskog razvoja. Embrionalna sličnost se sada objašnjava stvarnim odnosom organizama, a njihovo postepeno divergencija (embrionalna divergencija) služi kao očigledan odraz povijesne divergencije ovih oblika (filogenetska divergencija). U embrionu naših potomaka, napisao je Darwin, vidimo „nejasan portret“ naših predaka. Shodno tome, istorija date vrste može se pratiti individualnim razvojem.
Animal Biology
Sličnost između člankonožaca i anelida leži u karakteristikama kao što su segmentacija tijela, trbušna živčana vrpca i prisutnost cirkulacijskog sistema. Također srodan i potiče od drevnih anelida. Njihove razlike su...
Genetika i evoluciona nastava
Genetika i evoluciona nastava
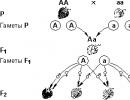
Ovaj zakon kaže da ukrštanje jedinki koje se razlikuju po datoj osobini (homozigotnih za različite alele) proizvodi genetski homogeno potomstvo (F 1 generacija), od kojih su sve jedinke heterozigotne...
Genetika i evolucija. Mendelovi zakoni genetike
Ovaj zakon se zove zakon (nezavisnog) cijepanja. Njegova suština je sljedeća. Kada organizam heterozigotan za ispitivanu osobinu razvije polne ćelije - gamete, tada jedna polovina njih nosi jedan alel datog gena...
Genetika i evolucija. Mendelovi zakoni genetike
Ovaj zakon kaže da se svaki par alternativnih karakteristika ponaša nezavisno jedna od druge u nizu generacija, usled čega među potomcima prve generacije (tj.
Ovaj zakon se zove zakon (nezavisnog) cijepanja. Njegova suština je sljedeća. Kada organizam heterozigotan za ispitivanu osobinu razvije polne ćelije - gamete, tada jedna polovina njih nosi jedan alel datog gena...
Genetika i evolucija. Osnovni aksiomi biologije
Ovaj zakon kaže da se svaki par alternativnih karakteristika ponaša nezavisno jedna od druge u nizu generacija, usled čega među potomcima prve generacije (tj.
Zakon radioaktivnog raspada
U gornjem matematičkom izrazu, to je konstanta raspada, koja karakteriše vjerovatnoću radioaktivnog raspada u jedinici vremena i ima dimenziju c?1. Znak minus označava smanjenje broja radioaktivnih jezgara tokom vremena...
Čovjekovo mjesto u prirodi
Tip Chordata: · skelet formira notohorda - visoko vakuolizirano vezivno tkivo...
Osnovni pojmovi savremene prirodne nauke
Određene vrste varijabilnosti su periodične. Oni su “vibracije” koje određuju nivoe bića u njihovoj hijerarhiji. Sve je u svemiru živo, ne računajući "periferiju" - donji rub skale "vodika". Odnosno...
Postanak i evolucija čovjeka
Prije nego što govorimo o vremenu pojave čovjeka, moramo razjasniti pitanje razlike između čovjeka i životinje, jer ideja o tome što je čovjek donosi zaključke o njegovom formiranju. Prvo, o sličnostima između ljudi i životinja...
Korelacija između onto- i filogenije. Zakon germinativne sličnosti
Biota epifitskih lišajeva mješovitih i listopadnih šuma u blizini sela Zilair, okrug Zilair
Obično se u tu svrhu koristi koeficijent sličnosti...
Godine 1828. Karl von Baer je formulirao obrazac: " Što se uporede raniji stadijumi individualnog razvoja, to se više može naći sličnosti Upoređujući faze razvoja embriona različitih vrsta i klasa hordata, K. Baer je donio sljedeće zaključke.
Embrioni životinja istog tipa su slični u ranim fazama razvoja.
Dosljedno se kreću u svom razvoju od opštijih karakteristika tipa ka sve specifičnijim. Konačno, razvijaju se znakovi koji ukazuju da embrij pripada određenom rodu, vrsti i, konačno, individualnim osobinama.
Embriji različitih predstavnika istog tipa postepeno se odvajaju jedan od drugog.
Baer je formulirao zakone germinativne sličnosti:
Najopštiji karakteri bilo koje velike grupe životinja pojavljuju se u embriju ranije od manje opštih karaktera;
Nakon formiranja najopštijih karakteristika javljaju se manje opšte, i tako sve do pojave posebnih karakteristika karakterističnih za datu grupu;
Embrion bilo koje vrste životinja, kako se razvija, postaje sve manje sličan embrionima drugih vrsta i ne prolazi kroz kasnije faze njihovog razvoja;
Zametak visoko organizirane vrste može nalikovati embriju primitivnije vrste, ali nikada nije sličan odraslom obliku ove vrste.
Sam Karl Baer nije prihvatio evolucijska učenja Charlesa Darwina, ali biolozi smatraju njegove zakone “embriološkim dokazom evolucije”.
“Na primjer, tokom devet mjeseci provedenih u maternici, ljudski embrion prolazi kroz mnoge faze - od beskičmenjaka do ribe, zatim do vodozemca, do reptila, sisara, primata, do sličnog homininu i čovjeka kao takvog.
Von Baerov zakon implicira da se evolucijske promjene češće dešavaju u kasnijim fazama razvoja, dok su rane faze evolucijski konzervativnije. To je zato što je vjerojatnije da će svaka mutacija koja utječe na rani razvoj proizvesti značajan fenotipski učinak nego ona koja utječe na kasni razvoj. Budući da je razvoj kontinuiran i kumulativan, promjene u ranoj fazi će imati sve veće posljedice od promjena u kasnijim fazama razvoja. Najvjerovatniji ishod bilo koje mutacije koja se dogodi u ranoj fazi je nepovoljan i često fatalan. Relativno kasnije (u odnosu na period razvoja) mutacije vjerojatnije neće imati negativne efekte, au nekim slučajevima mogu čak i povećati adaptaciju kroz suptilne promjene fenotipa.( Ovaj fenomen se može ilustrovati povlačenjem analogije sa izgradnjom nebodera. Ako se naprave promjene u dizajnu zidova prizemlja, postoji velika šansa da će svaki sprat iznad biti pogođen, moguće na negativan način. Bilo kakve promjene na gornjem spratu nebodera neće uticati na niže spratove.)
Von Baerov zakon više vrijedi za organizme koji se razvijaju unutar majke (kao što su sisari) nego za vrste koje imaju stadij larve u kojem se moraju sami brinuti.
Tokom intrauterinog razvoja, pritisak prirodne selekcije iz vanjskog okruženja koji dovodi do promjena je minimalan ili ga nema. Međutim, organizam larve, koji samostalno osigurava svoj opstanak, stalno je podložan pritisku prirodne selekcije. Ovo objašnjava zašto su rani razvojni stadijumi sisara toliko slični među vrstama, ali zašto se faza larve organizama kao što su insekti veoma razlikuje od faze odrasle osobe.”
U kojim fazama embriogeneze nastaju blastokoel, gastrocel i celim?
Faza drobljenja. Blastocoel- šupljina blastule nastala 4-5 dana cijepanja između blastomera u životinjskim embrionima. Napunjen tečnošću koja se po hemijskom sastavu razlikuje od okoline. Šupljina povećava površinu embrija, poboljšavajući njegovu sposobnost da apsorbira hranjive tvari i kisik. Najveću veličinu dostiže pri kraju cijepanja, u fazi blastule. Tokom procesa gastrulacije, postepeno se istiskuje tokom invaginacije (invaginacije) embrionalnog zida ili se puni (u toku imigracije) pokretnim ćelijama.
Faza gastrulacije. Gastrocoel- primarno crijevo, šupljina gastrule, koja nastaje kod višećelijskih embrija ako se gastrulacija provodi intususcepcijom. Gastrocoel je ispunjen tečnošću i komunicira sa spoljašnjim okruženjem kroz poseban otvor - blastopore. Zidovi gastrocela sastoje se od invaginirajućeg primarnog endoderma. Nakon toga, gastrocoel postaje šupljina konačnog crijeva.
Faza diferencijacije mezoderma. Sveukupno- sekundarna tjelesna šupljina višećelijskih životinja. U trohoforima se formira od specijalizovanih mezodermalnih ćelija - teloblasta kao rezultat njihove deobe i naknadnog formiranja šupljina unutar nastalih grupa ćelija. Ova metoda formiranja celima u ontogenezi se naziva teloblastična. Kod deuterostoma, celim nastaje izbočenjem zidova primarnog crijeva i odvajanjem rezultirajućih izbočina. Ova metoda formiranja celima naziva se enterokoelozna. U oba slučaja, celim se smatra mezodermalnom formacijom. Razlikuje se od primarne tjelesne šupljine po prisutnosti vlastite epitelne obloge (zida). Epitel koji formira oblogu celima naziva se celiotel ili mezotel.
Razdvajanje. U roku od 3-4 dana dolazi do drobljenja u jajovodu. Light mali blastomeri brže se fragmentiraju i okružuju mračno velike koje ostaju unutra. Embrion bez šupljine, koji se sastoji od guste nakupine blastomera, naziva se Morula( nastaje 3-4. dana drobljenja). 4-5 dana embrion ulazi u šupljinu materice, odakle apsorbuje tečnost i akumulira je u svojoj šupljini - blastocoel . Njegov zid formiraju mali, lagani blastomeri - trofoblast . Tamne ćelije se potiskuju na jedan od polova i formiraju embrioblast . Rezultat blastula pozvao blastocista ili blastodermične vezikule. Do 7 dana blastocista ostaje u šupljini materice u slobodnom stanju, hraneći se sekretom žlijezda materice. Ovim se završava početni (1. sedmica) period embriogeneze
Intususcepcija. Uočava se kod životinja s izolecitalnim tipom jaja (holoturija, kopljasta). Vegetativni pol blastule invaginira prema unutra. Kao rezultat toga, suprotni polovi blastoderma se praktično zbližavaju, tako da blastokoel ili nestaje ili ostaje mali jaz. Kao rezultat toga nastaje dvoslojni embrion čiji je vanjski zid primarni ektoderm, a unutrašnji zid primarni endoderm.Invaginacija formira primarni crijevo-arhenteron ili gastrocoel.Otvor kroz koji komunicira sa spoljašnje okruženje. nazvana primarna usta ili blastopore.
Diferencijacija mezoderma. Ventralni dijelovi mezoderma nisu segmentirani i formiraju splanhnotom.Podijeljen je na dva sloja - parijetalni i visceralni, koji okružuju sekundarnu tjelesnu šupljinu - coelom.
GASTRULACIJA
Suština faze gastrulacije leži u činjenici da se jednoslojni embrion - blastula - pretvara u višeslojni - dvo- ili troslojni, tzv gastrula(iz grčkog gaster - stomak u umanjenom značenju).
Kod primitivnih hordata, na primjer, lanceta, homogeni jednoslojni blastoderm tokom gastrulacije se transformira u vanjski zametni sloj - ektoderm - i unutrašnji zametni sloj - endoderm. Endoderm formira primarno crijevo sa šupljinom unutar gastrocele. Rupa koja vodi u gastrocoel naziva se blastopore ili primarna usta. Dva klica su morfološki znaci gastrulacije. Njihovo postojanje na određenom stupnju razvoja kod svih višećelijskih životinja, od koelenterata do viših kralježnjaka, omogućava nam da razmišljamo o homologiji zametnih slojeva i jedinstvu porijekla svih ovih životinja.
Kod kičmenjaka, pored dva pomenuta tokom gastrulacije, formira se i treći zametni sloj - mezoderm, zauzimaju mjesto između ekto- i endoderme. Razvoj srednjeg zametnog sloja, koji je hordomezoderm, evolucijska je komplikacija faze gastrulacije u kralježnjaka i povezana je s ubrzanjem njihovog razvoja u ranim fazama embriogeneze. Kod primitivnijih hordata, kao što je lanceta, hordomezoderma se obično formira na početku sljedeće faze nakon gastrulacije - organogeneza. Pomak u vremenu razvoja nekih organa u odnosu na druge kod potomaka u odnosu na grupe predaka je manifestacija heterohronija. Promjene u vremenu formiranja najvažnijih organa u procesu evolucije nisu neuobičajene.
Karakteriziran je proces gastrulacije važne stanične transformacije, kao što su usmjerena kretanja grupa i pojedinačnih stanica, selektivna proliferacija i sortiranje stanica, početak citodiferencijacije i induktivne interakcije.
Metode gastrulacije su različiti. Postoje četiri vrste prostorno usmjerenih kretanja ćelija koje dovode do transformacije embrija iz jednoslojnog u višeslojni.
Intususcepcija - invaginacija jednog od dijelova blastoderma prema unutra kao cijeli sloj. Kod lancete invaginiraju ćelije vegetativnog pola, a kod vodozemaca invaginacija se javlja na granici između životinjskog i vegetativnog pola u području sivog sokola. Proces invaginacije moguć je samo u jajima sa malom ili srednjom količinom žumanca.
Epiboly - prerast malih ćelija životinjskog pola sa većim ćelijama vegetativnog pola koje zaostaju u brzini deobe i manje su pokretljive. Ovaj proces je jasno izražen kod vodozemaca.
Denominacija - razdvajanje ćelija blastoderma u dva sloja koji leže jedan iznad drugog. Delaminacija se može uočiti u diskoblastuli embriona s djelomičnim tipom cijepanja, kao što su gmizavci, ptice i sisari koji su oviparali. Delaminacija se javlja u embrioblastu placentnih sisara, što dovodi do stvaranja hipoblasta i epiblasta.
imigracija - kretanje grupa ili pojedinačnih ćelija koje nisu ujedinjene u jedan sloj. Imigracija se javlja kod svih embrija, ali je najkarakterističnija za drugu fazu gastrulacije kod viših kralježnjaka.
U svakom konkretnom slučaju embriogeneze, u pravilu se kombinira nekoliko metoda gastrulacije.
Karakteristike faze gastrulacije. Gastrulaciju karakteriziraju različiti ćelijski procesi. Mitotić nastavlja proliferacija ćelija,Štaviše, ima različit intenzitet u različitim dijelovima embrija. Međutim, najkarakterističnija karakteristika gastrulacije je kretanje ćelijskih masa. To dovodi do promjene strukture embrija i njegove transformacije iz blastule u gastrulu. Dešava se sortiranjećelije prema njihovoj pripadnosti različitim zametnim slojevima, unutar kojih se međusobno „prepoznaju“.
Počinje faza gastrulacije citodiferencijacija,što znači prijelaz na aktivno korištenje bioloških informacija iz vlastitog genoma. Jedan od regulatora genetske aktivnosti je različit hemijski sastav citoplazme embrionalnih ćelija, nastao kao rezultat ovoplazmatske segregacije. Tako su ektodermalne ćelije vodozemaca tamne boje zbog pigmenta koji je u njih ušao iz životinjskog pola jajeta, a ćelije endoderme su svijetle, budući da potiču iz vegetativnog pola jajeta.
Tokom gastrulacije, uloga embrionalna indukcija. Pokazalo se da je pojava primitivne pruge kod ptica rezultat induktivne interakcije između hipoblasta i epiblasta. Hipoblast karakteriše polaritet. Promjena položaja hipoblasta u odnosu na epiblast uzrokuje promjenu orijentacije primitivne pruge.
Formiranje zigote. Razdvajanje. Vrste drobljenja.
Zigota- diploidna ćelija nastala kao rezultat fuzije haploidnih ženskih i muških gameta (oplođeno jaje).
razdvajanje - kontinuirana mitotska dioba diploidnih stanica (zigota i blastomera) bez povećanja njihovog ukupnog volumena.
Formiranje zigote.
Nakon spajanja vanjskih membrana, jezgro spermatozoida se nalazi unutar jajeta. Od ovog trenutka, jezgra jajne ćelije i sperme nazivaju se ženskim i muškim pronukleusima. Muški pronukleus se okreće tako da su centriole bliže centru jajeta i počinju se kretati dublje u citoplazmu. Tokom ovog perioda, DNK se udvostručuje u oba pronukleusa. Nakon toga, morski jež prolazi kroz fuziju pronukleusa i formira diploidno jezgro zigote. Kod većine životinja i ljudi, svaki pronukleus prolazi kroz profazu mitoze, a njihove nuklearne membrane se raspadaju. Zatim se hromozomski skupovi pronukleusa kombinuju, a rezultirajući diploidni skup formira metafaznu ploču. Odmah nakon toga u zigoti se javlja prva mitoza i citokineza.
Cijepanje se razlikuje od normalnog ćelijskog ciklusa po odsustvu G1 i G2 faza i vrlo kratkoj S fazi. Interval između podjela drobljenja je otprilike 12-24 sata. Priroda fragmentacije i rezultirajuća vrsta blastule određena je brojem i distribucijom inkluzija žumanjka u citoplazmi jajeta.
Vrste drobljenja:
· Holoblastični tip drobljenja(potpuna ujednačena fragmentacija) karakteristična je za izolecitalno jaje. Zigota i blastomeri su potpuno podijeljeni na ćelije kćeri jednake veličine. Kod sinhronog tipa fragmentacije uočava se geometrijska progresija, uzastopno se formira 2,4,8,16,32,64,128 blastomera, a kod asinhronog tipa formira se 3-ćelijska faza.
· Potpuno neravnomjerno drobljenje nastaje u oplođenom telolecitalnom jajetu, jaje se u potpunosti dijeli, na životinjskom polu dolazi do bržeg cijepanja i formiraju se brojne male ćelije, a na vegetativnom polu lokaliziraju se velike stanice opterećene žumanjkom.
· Meroblastični tip drobljenja (nepotpuno, djelomično drobljenje) javlja se u oštro oplođenim telolecitalnim i centrolecitalnim jajima, citoplazma bez žumanca se drobi.
· Diskoidno nepotpuno drobljenje karakterizira fragmentacija citoplazme koja sadrži jezgro u području životinjskog pola.
Po vrsti simetrije:
· Radijalno(os jajne ćelije je os radijalne simetrije blastomera koji se dijele)
· Spiralna(blastomeri koji nastaju naknadnim podjelama nalaze se jedan u odnosu na drugi u spiralu)
· Bilateralno( blastomeri su raspoređeni na takav način da embrion ima samo jednu ravan simetrije)
· anarhičan ( kod jedne vrste postoje različite opcije za lokaciju blastomera - lanci ili bezoblična ćelijska masa)
Ontogeneza – implementacija genetske informacije koja se javlja u svim fazama.
Ontogeneza je genetski kontrolisan proces. Tokom ontogeneze ostvaruje se genotip i formira fenotip.
Ontogeneza je individualni razvoj organizma, skup uzastopnih morfoloških, fizioloških i biohemijskih transformacija kroz koje organizam prolazi od trenutka svog nastanka do kraja života. O. uključuje visina odnosno povećanje telesne težine, njene veličine, diferencijaciju. Izraz "O." uveo E. Haeckel(1866) kada je formulisao biogenetski zakon.
Prvi pokušaj istorijskog potkrepljenja O. napravio je I. f. Meckel. Problem odnosa između O. i filogenije postavio je Ch. Darwin a razvio ga je F. Muller,E. Haeckel i dr. U O. nastaju svi evolucijski novi karakteri povezani s promjenama u nasljeđu, ali se pri tome čuvaju samo oni koji doprinose boljoj adaptaciji organizma na uslove postojanja. prirodna selekcija i prenose se na naredne generacije, tj. fiksiraju se u evoluciji. Poznavanje zakona, uzroka i faktora kiseonika služi kao naučna osnova za pronalaženje sredstava uticaja na razvoj biljaka, životinja i ljudi, što je od izuzetne važnosti za biljnu i stočarsku proizvodnju, kao i za medicinu. .
Filogenija je istorijski razvoj organizama. Termin je uveden u njemački jezik. evolucionista E. Haeckel 1866. Glavni zadatak u proučavanju fiziologije je rekonstruisati evolucionu transformaciju životinja, biljaka i mikroorganizama i na osnovu toga utvrditi njihovo porijeklo i srodne odnose između svojti kojima proučavani organizmi pripadaju. U tu svrhu, E. Haeckel je razvio metodu „trostrukog paralelizma“, koja omogućava da se upoređivanjem podataka iz tri nauke — morfologije, embriologije i paleontologije — rekonstruiše tok istorijskog razvoja sistematske grupe koja se proučava.
Zakon germinativne sličnosti
Istraživači ranog 19. veka. po prvi put su počeli obraćati pažnju na sličnost faza razvoja embriona viših životinja sa fazama složenosti organizacije koje vode od nisko organiziranih oblika ka progresivnim. Upoređujući faze razvoja embriona različitih vrsta i klasa hordata, K. Baer je došao do sljedećih zaključaka.
1. Embrioni životinja istog tipa su slični u ranim fazama razvoja.
2. Dosljedno se kreću u svom razvoju od opštijih karakteristika tipa ka sve specifičnijim. Konačno, razvijaju se znakovi koji ukazuju da embrij pripada određenom rodu, vrsti i, konačno, individualnim osobinama.
3. Embrioni različitih predstavnika istog tipa postepeno se odvajaju jedan od drugog.
K. Baer, budući da nije evolucionista, nije mogao povezati obrasce individualnog razvoja koje je otkrio s procesom filogeneze. Stoga su generalizacije koje je napravio imale značenje samo empirijskih pravila.
Naknadni razvoj evolucijske ideje omogućio je da se sličnost ranih embriona objasni njihovim istorijskim srodstvom, i njihovo sticanje sve više posebnih osobina uz postepenu izolaciju jedna od druge - stvarna izolacija odgovarajućih klasa, redova, porodica, rodova i vrsta u procesu evolucije.
Ubrzo nakon otkrića zakona embrionalne sličnosti, Charles Darwin je pokazao da ovaj zakon svjedoči o zajedničkom porijeklu i jedinstvu početnih faza evolucije unutar tipa.
Biogenetski zakon Haeckel-Müller: svako živo biće u svom individualnom razvoju ( ontogeneza) u određenoj mjeri ponavlja forme kroz koje su prolazili njegovi preci ili njegova vrsta ( filogeneza).
Ontogeneza - ponavljanje filogeneze
Uspoređujući ontogenezu rakova sa morfologijom njihovih izumrlih predaka, F. Muller je zaključio da živi rakovi u svom razvoju ponavljaju put koji su prošli njihovi preci. Transformacija ontogeneze u evoluciji, prema F. Mulleru, vrši se zbog njenog produžavanja dodavanjem dodatnih faza ili proširenja. Na osnovu ovih zapažanja, kao i proučavanja razvoja hordata, E. Haeckel (1866) je formulisao osnovni biogenetički zakon, prema kojem je Ontogeneza je kratko i brzo ponavljanje filogenije.
Ponavljanje struktura karakterističnih za pretke u embriogenezi potomaka naziva se rekapitulacije. Rekapituliraju se ne samo morfološke karakteristike notohorda, lanca škržnih proreza i škržnih lukova kod svih horda, već i karakteristike biohemijske organizacije i fiziologije. Dakle, u evoluciji kralježnjaka dolazi do postepenog gubitka enzima neophodnih za razgradnju mokraćne kiseline, produkta metabolizma purina. Kod većine beskičmenjaka krajnji produkt razgradnje mokraćne kiseline je amonijak, kod vodozemaca i riba to je urea, kod mnogih gmizavaca alantoin, a kod nekih sisara mokraćna kiselina se uopće ne razgrađuje i izlučuje se urinom. U embriogenezi sisara i ljudi bilježe se biohemijske i fiziološke rekapitulacije: oslobađanje amonijaka od ranih embrija, kasnije uree, zatim alantoina, a u posljednjim fazama razvoja - mokraćne kiseline.
Međutim, u ontogenezi visokoorganiziranih organizama ne opaža se uvijek striktno ponavljanje faza historijskog razvoja, kao što slijedi iz biogenetskog zakona. Dakle, ljudski embrion nikada ne ponavlja odrasle stadijume riba, vodozemaca, gmizavaca i sisara, već je po nizu osobina sličan samo njihovim embrionima. Rane faze razvoja ostaju najkonzervativnije, zbog čega se potpunije rekapituliraju od kasnijih. To je zbog činjenice da je jedan od najvažnijih mehanizama za integraciju ranih faza embriogeneze embrionalna indukcija, a strukture embrija koje se prve formiraju, kao što su notohorda, neuralna cijev, ždrijelo, crijeva i somiti, predstavljaju organizacione centre embrija, od kojih zavisi ceo tok razvoja.
Genetska osnova rekapitulacije leži u jedinstvu mehanizama genetičke kontrole razvoja, koji se čuva na osnovu zajedničkih gena za regulaciju ontogeneze, koje nasljeđuju srodne grupe organizama od zajedničkih predaka.
Rekapitulacija(od latinskog recapitulatio - ponavljanje) - koncept koji se koristi u biologiji za označavanje ponavljanja u individualnom razvoju karakteristika karakterističnih za raniju fazu evolucijskog razvoja.
Ontogeneza kao osnova filogeneze. Cenogeneza. Autonomija ontogenije. Filembriogeneza. Doktrina A.N. Severtsova o filebriogenezi. Mehanizmi njihovog nastanka. Heterohronije i heterotopije bioloških struktura u evoluciji ontogeneze.
Oslanjajući se samo na osnovni biogenetički zakon, nemoguće je objasniti proces evolucije: beskonačno ponavljanje onoga što je prošlo samo po sebi ne rađa nešto novo. Budući da život na Zemlji postoji zahvaljujući smjeni generacija određenih organizama, njegova evolucija nastaje zbog promjena koje se dešavaju u njihovoj ontogeniji. Ove promjene se svode na činjenicu da specifične ontogenije odstupaju od puta koji su zacrtali oblici predaka i dobijaju nove karakteristike.
Takva odstupanja uključuju npr. cenogeneza - adaptacije koje nastaju u embrionima ili larvama i prilagođavaju ih karakteristikama njihovog okruženja. Kod odraslih organizama cenogeneza nije očuvana. Primjeri koenogeneze su rožnate formacije u ustima ličinki vodozemaca bez repa, koje im olakšavaju hranu biljnom hranom. Tokom procesa metamorfoze u žabi, oni nestaju i probavni sistem se obnavlja da se hrani insektima i crvima. Koenogeneza kod amniota uključuje embrionalne membrane, žumančanu vreću i alantois, a kod placentnih sisara i ljudi i placentu i pupčanu vrpcu.
Cenogeneza, koja se pojavljuje tek u ranim fazama ontogeneze, ne mijenja tip organizacije odraslog organizma, ali daje veću vjerovatnoću preživljavanja potomstva. Oni mogu biti praćeni smanjenjem plodnosti i produženjem embrionalnog ili larvalnog perioda, zbog čega se organizam u postembrionalnom ili postlarvalnom razdoblju razvoja pokazuje zrelijim i aktivnijim. Nakon što su nastale i pokazale se korisnim, cenogeneze će se razmnožavati u narednim generacijama. Dakle, amnion, koji se prvi put pojavio kod predaka gmizavaca u karbonskom periodu paleozojske ere, reproducira se kod svih kralježnjaka koji se razvijaju na kopnu, kako kod jajonosnih reptila i ptica, tako i kod placentnih sisara.
Druga vrsta filogenetski značajnih transformacija filogenije je filebriogeneza. Predstavljaju odstupanja od ontogeneze karakteristična za pretke, koja se manifestuju u embriogenezi, ali imaju adaptivni značaj kod odraslih oblika. Tako se pupoljci dlake pojavljuju kod sisara u vrlo ranim fazama embrionalnog razvoja, ali sama dlaka je važna samo kod odraslih organizama.
Takve promjene u ontogenezi, budući da su korisne, fiksiraju se prirodnom selekcijom i reprodukuju u narednim generacijama. Ove promjene se zasnivaju na istim mehanizmima koji uzrokuju kongenitalne malformacije: poremećaj proliferacije ćelija, njihovo kretanje, adhezija, smrt ili diferencijacija (vidi § 8.2 i 9.3), međutim, kao i cenogeneza, razlikuju se od defekata po adaptivnoj vrijednosti, tj. korisnost i fiksacija prirodnom selekcijom u filogenezi.
U zavisnosti od faza embriogeneze i morfogeneze određenih struktura, nastaju razvojne promene koje imaju značaj filebriogeneze, razlikuju se tri tipa.
1.anabolizam, ili ekstenzije, nastaju nakon što je organ praktično završio svoj razvoj, a izražavaju se u dodavanju dodatnih faza koje mijenjaju konačni rezultat.
Anabolizmi uključuju fenomene kao što je iverak sticanje specifičnog oblika tijela tek nakon što se iz jajeta izleže mladica, koja se ne razlikuje od ostalih riba, kao i pojavu zakrivljenosti kralježnice, spajanje šavova u moždanoj lubanji i konačno preraspodjela krvnih žila u tijelu sisara i ljudi.
2.odstupanja - devijacije koje nastaju u procesu morfogeneze organa. Primjer je razvoj srca u ontogenezi sisara, u kojem ono rekapitulira stadij cijevi, dvokomornu i trokomornu strukturu, ali fazu formiranja nepotpunog septuma, karakterističnog za gmizavce, zamjenjuje razvojem pregrade, građene i locirane drugačije i karakteristične samo za sisare (videti § 14.4). Razvoj pluća kod sisara takođe otkriva rekapitulaciju ranih faza predaka; kasnija morfogeneza se odvija na nov način (videti odeljak 14.3.4 ).
Rice. 13.9. Transformacije onto- i filogeneze u vezi sa novonastalom filebriogenezom
Slova označavaju faze ontogeneze, brojevi označavaju filebriogenetske transformacije
3.arhalaksija - promjene koje se otkrivaju na nivou primordija i izražavaju se u kršenju njihove podjele, ranoj diferencijaciji ili u pojavi fundamentalno novih primordija. Klasičan primjer arhalaksije je
razvoj dlake kod sisara, čija se bradavica javlja u vrlo ranim fazama razvoja i od samog početka razlikuje se od dlaka drugih kožnih dodataka kičmenjaka (vidi § 14.1).
Prema tipu arhalaksije, notohorda se pojavljuje kod primitivnih riba bez lobanje, hrskavična kralježnica kod hrskavičnih riba (vidi odjeljak 14.2.1.1), a nefroni sekundarnog bubrega razvijaju se kod gmizavaca (vidi odjeljak 14.5.1).
Jasno je da se tokom evolucije zbog anabolizma u ontogenijima potomaka u potpunosti ostvaruje osnovni biogenetski zakon, tj. dešavaju se rekapitulacije svih faza razvoja predaka. U slučaju odstupanja, rani stadijumi predaka se rekapituliraju, a kasnije se zamjenjuju razvojem u novom smjeru. Arhalaksija u potpunosti sprečava rekapitulaciju u razvoju ovih struktura, mijenjajući same njihove rudimente.
Ako uporedimo dijagram filebriogeneze sa tabelom K. Baera (slika 13.9), koja ilustruje zakon germinalne sličnosti, postaje jasno da je Baer već bio veoma blizu otkriću filebriogeneze, ali odsustvo evolucione ideje u njegovom rasuđivanju nije dozvolio da bude više od 100 godina ispred naučne misli .
U evoluciji ontogeneze anabolizam se najčešće javlja kao filebriogeneza, koja samo u maloj mjeri mijenja cjelokupni razvojni proces. Odstupanja kao kršenja morfogenetskog procesa u embriogenezi često se odbacuju prirodnom selekcijom i stoga su mnogo rjeđa. Arhalaksija se najrjeđe javlja u evoluciji zbog činjenice da mijenjaju cijeli tok embriogeneze, a ako takve promjene zahvate rudimente vitalnih organa ili organa koji su embrionalni organizacioni centri (vidi odjeljak 8.2.6), onda se često ispadaju kao biti nespojivo sa životom.
U istoj filogenetskoj grupi, evolucija u različitim sistemima organa može se dogoditi zbog različite filebriogeneze.
Tako se u ontogenezi sisara prate svi stadijumi razvoja aksijalnog skeleta u podtipu kičmenjaka (anabolizam), u razvoju srca se rekapituliraju samo rani stadijumi (devijacija), a u razvoju kožnih privjesaka postoji nema rekapitulacije uopšte (arhalaksija). Poznavanje tipova filebriogeneze u evoluciji organskih sistema hordata je neophodno da bi lekar predvideo mogućnost pojave kongenitalnih malformacija atavističke prirode kod fetusa i novorođenčadi (videti odeljak 13.3.4). su mogući u organskom sistemu koji se razvija kroz anabolizam i devijacije zbog rekapitulacije stanja predaka, onda je u slučaju arhalaksije to potpuno isključeno.
Osim cenogeneze i filebriogeneze, odstupanja u vremenu formiranja organa mogu se naći i u evoluciji ontogeneze - heterohronija - i mesta njihovog razvoja - heterotopija. I prvi i drugi dovode do promjena u međusobnoj korespondenciji struktura u razvoju i podliježu strogoj kontroli prirodne selekcije. Sačuvane su samo one heterohronije i heterotopije koje se pokažu korisnima. Primjeri takvih adaptivnih heterohronija su pomaci u vremenu formiranja najvitalnijih organa u grupama koje se razvijaju prema tipu arogeneze. Dakle, kod sisara, a posebno kod ljudi, diferencijacija prednjeg mozga značajno pospješuje razvoj ostalih njegovih dijelova.
Heterotopije dovode do stvaranja novih prostornih i funkcionalnih veza između organa, osiguravajući njihovu daljnju zajedničku evoluciju. Tako srce, koje se nalazi ispod ždrijela ribe, osigurava efikasan protok krvi u škržne arterije za razmjenu plinova. Prelazeći u retrosternalnu regiju kod kopnenih kralježnjaka, razvija se i funkcionira u jednom kompleksu s novim respiratornim organima - plućima, i ovdje, prije svega, obavlja funkciju isporuke krvi u respiratorni sistem za izmjenu plinova.
Heterohronije i heterotopije, u zavisnosti od toga u kojim fazama embriogeneze i morfogeneze organa se pojavljuju, mogu se smatrati filebriogenezom različitih tipova. Dakle, kretanje rudimenata mozga, koje dovodi do njegovog savijanja, karakteristično za amniote, a koje se manifestuje u početnim fazama njegove diferencijacije, je arhalaksija i heterotopija testisa kod ljudi od trbušne šupljine kroz ingvinalni kanal do skrotum, uočen na kraju embriogeneze nakon njegovog konačnog formiranja, - tipična anabolija.
Ponekad procesi heterotopije, identični u rezultatima, mogu biti filebriogeneza različitih tipova. Na primjer, u različitim klasama kralježnjaka, pomicanje pojasa udova je vrlo uobičajeno. U mnogim grupama riba koje vode način života na dnu, trbušna peraja (zadnji udovi) nalaze se ispred prsnih peraja, a kod sisara i ljudi rameni pojas i prednji udovi u konačnom stanju nalaze se znatno kaudalno od mjesta njihovo početno formiranje. S tim u vezi, inervaciju ramenog pojasa u njima provode živci povezani ne s torakalnim, već s cervikalnim segmentima kičmene moždine. Kod gore navedenih riba, karlične peraje inerviraju ne zadnji nervi trupa, već prednji segmenti, koji se nalaze ispred centara inervacije prsnih peraja. To ukazuje na heterotopiju u formiranju peraja već u fazi najranijih rudimenata, dok se pomicanje prednjeg pojasa udova kod ljudi javlja u kasnijim fazama, kada je njihova inervacija već u potpunosti ostvarena. Očigledno, u prvom slučaju heterotopija predstavlja arhalaksu, au drugom anabolizam.
Cenogeneza, filebriogeneza, kao i heterotopija i heterohronija, koji su se pokazali korisnima, fiksiraju se u potomstvu i reprodukuju u narednim generacijama sve dok ih nove adaptivne promjene u ontogenezi ne istisnu i zamijene. Zahvaljujući tome, ontogeneza ne samo da nakratko ponavlja evolucijski put koji su prošli naši preci, već i utire nove pravce filogeneze u budućnosti.
Cenogeneza
(od grčkog kainós - novi i ... geneza (Vidi ... geneza)
adaptacija organizma koja se javlja u embrionalnom (fetalnom) ili larvalnom stadiju i ne perzistira u odrasloj jedinki. Primeri c. su placenta sisara, koja fetusu obezbeđuje disanje, ishranu i izlučivanje; vanjske škrge larvi vodozemaca; zub jajeta kod ptica, koji pilići koriste da probiju ljusku jajeta; organi vezivanja u larvi ascidijana, plivajući rep u larvi trematoda - cercariae, itd. Izraz "C." koju je 1866. uveo E. Haeckel da označi one karakteristike koje, narušavajući manifestacije palingeneze (vidi. Palingeneza), tj. ponavljanja udaljenih faza filogeneze u procesu embrionalnog razvoja pojedinca, ne dozvoljavaju nam da pratimo slijed faza filogeneze njihovih predaka tokom ontogeneze modernih oblika, tj. krše Biogenetski zakon. Krajem 19. vijeka. Svaka promjena u toku ontogeneze karakteristična za pretke počela je da se naziva bojom (njemački naučnici E. Mehnert, F. Keibel i drugi). Moderno razumijevanje pojma "C." nastao je kao rezultat rada A. N. Severtsova, koji je za ovaj koncept zadržao samo značenje privremenih uređaja, odnosno embrionalnih adaptacija. vidi takođe Filembriogeneza.
Cenogeneza(grč. kainos novi + geneza porijeklo, formiranje) - pojava u embrionu ili larvi adaptacija na uslove života koji nisu karakteristični za odrasle faze, na primjer. formiranje membrana u embrionima viših životinja.
Filembriogeneza
(od grčkog phýlon - pleme, rod, vrsta i embriogeneza
FILEMBRIOGENEZA (od grčkog phylon - rod, pleme, embrion - embrion i geneza - porijeklo), evolucijska promjena ontogenija organa, tkiva i ćelija, povezanih s progresivnim razvojem i smanjenjem. Doktrinu o filembriogenezi razvio je ruski evolucijski biolog A.N. Severtsov. Načini (metode) filebriogeneze razlikuju se po vremenu nastanka tokom razvoja ovih struktura.
Ako se razvoj određenog organa kod potomaka nastavi nakon faze na kojoj je završio kod predaka, dolazi do anabolije (od grčkog anabole - uzdizanje) - produžetka završne faze razvoja. Primjer je formiranje srca sa četiri komore kod sisara. Vodozemci imaju trokomorno srce: dva atrija i jednu komoru. Kod gmizavaca se u komori razvija septum (prva anabolija), ali kod većine je ovaj septum nekompletan – samo smanjuje miješanje arterijske i venske krvi. Kod krokodila i sisara razvoj septuma se nastavlja do potpunog odvajanja desne i lijeve komore (druga anabolija). Kod djece je ponekad, kao atavizam, interventrikularni septum nedovoljno razvijen, što dovodi do ozbiljne bolesti koja zahtijeva hiruršku intervenciju.
Produženje razvoja organa ne zahtijeva duboke promjene u prethodnim fazama njegove ontogeneze, stoga je anabolizam najčešći metod filebriogeneze. Faze razvoja organa koje prethode anabolizmu ostaju uporedive sa fazama filogenija preci (tj. oni su rekapitulacije) i može poslužiti za njegovu rekonstrukciju (vidi. Biogenetski zakon). Ako razvoj organa u srednjim fazama odstupi od puta kojim se odvijala njegova ontogeneza kod njegovih predaka, dolazi do devijacije (od kasnolat. deviatio - odstupanje). Na primjer, kod riba i gmizavaca ljuske se pojavljuju kao zadebljanja epiderme i donjeg sloja vezivnog tkiva kože - korijuma. Postepeno se zgušnjavajući, ovaj anlage se savija prema van. Zatim kod riba korij okoštava, formirane koštane ljuske probijaju epidermu i pomiču se na površinu tijela. Kod gmizavaca, naprotiv, kost se ne formira, ali epiderma postaje keratinizirana, formirajući rožnate ljuske guštera i zmija. Kod krokodila, korij može okoštati, formirajući koštanu osnovu rožnatih ljuski. Odstupanja dovode do dubljeg restrukturiranja ontogeneze od anabolizma, pa su rjeđa.
Najrjeđe se javljaju promjene u primarnim rudimentima organa - archallaxis (od grčkog arche - početak i allaxis - promjena). U slučaju odstupanja, rekapitulacija se može pratiti od nastanka organa do trenutka razvojne devijacije. Kod arhalaksije nema rekapitulacije. Primjer je razvoj tijela kralježaka kod vodozemaca. Kod fosilnih vodozemaca - stegocefalija i kod modernih vodozemaca bez repa, tijela pršljenova su formirana oko tetive od nekoliko, obično po tri na svakoj strani tijela, odvojenih lanaca, koji se zatim spajaju u tijelo pršljena. Kod repatih vodozemaca ovi anlage se ne pojavljuju. Osifikacija raste iznad i ispod, pokrivajući notohordu, tako da se odmah formira koštana cijev, koja, zadebljajući, postaje tijelo pršljena. Ova arhalaksija je razlog za pitanje porijekla repatih vodozemaca o kojem se još uvijek raspravlja. Neki znanstvenici vjeruju da potječu direktno od riba s režnjevima, bez obzira na druge kopnene kralježnjake. Drugi kažu da su se repasti vodozemci vrlo rano odvojili od drugih vodozemaca. Drugi pak, zanemarujući razvoj pršljenova, dokazuju blisku vezu repatih i bezrepih vodozemaca.
Redukcija organa, koji su izgubili svoj adaptivni značaj, nastaju i kroz filebriogenezu, uglavnom kroz negativni anabolizam - gubitak završnih faza razvoja. U ovom slučaju, organ se ili nedovoljno razvija i postaje ostatak, ili podliježe obrnutom razvoju i potpuno nestaje. Primjer rudimenta je ljudsko slijepo crijevo - nerazvijeni cekum; primjer potpunog nestanka je rep punoglavaca žabe. Tokom svog života u vodi, rep raste, na njegovom kraju se dodaju novi pršljenovi i mišićni segmenti. Tokom metamorfoze, kada se punoglavac pretvori u žabu, rep se rastvara, a proces se odvija obrnutim redoslijedom - od kraja do baze. Filembriogeneza je glavna metoda adaptivnih promjena u strukturi organizama tokom filogeneze.
Principi (metode) filogenetskih transformacija organa i funkcija. Podudarnost strukture i funkcije u živim sistemima. Multifunkcionalnost. Kvantitativne i kvalitativne promjene u funkcijama bioloških struktura.
OPĆE REGULARNOSTI
EVOLUCIJA ORGANA
Organizam, ili pojedinac, je zasebno živo biće koje u procesu ontogeneze ispoljava sva svojstva živog bića. Stalna interakcija pojedinca sa okolinom u vidu organizovanih tokova energije i materije održava njen integritet i razvoj. Strukturno, tijelo je integrirani hijerarhijski sistem izgrađen od ćelija, tkiva, organa i sistema koji osiguravaju njegove vitalne funkcije. Pogledajmo bliže organe i sisteme za održavanje života.
Autoritet nazivaju istorijski razvijeni specijalizovani sistem tkiva, koji karakteriše razgraničenje, konstantnost oblika, lokalizacija, unutrašnja struktura cirkulatornih i inervacionih puteva, razvoj u ontogenezi i specifične funkcije. Struktura organa je često veoma složena. Većina njih je multifunkcionalna, tj. obavlja nekoliko funkcija istovremeno. Istovremeno, različiti organi mogu učestvovati u realizaciji bilo koje složene funkcije.
Skupina organa sličnog porijekla koji se kombinuju radi obavljanja složene funkcije naziva se sistem(cirkulacijski, ekskretorni, itd.).
Ako grupa organa različitog porijekla obavlja istu funkciju, naziva se aparata. Primjer je respiratorni aparat koji se sastoji od samih respiratornih organa i elemenata skeleta i mišićnog sistema koji obezbjeđuju respiratorne pokrete.
U procesu ontogeneze dolazi do razvoja i često zamjene nekih organa drugim. Organi zrelog organizma nazivaju se definitivan; organi koji se razvijaju i funkcionišu samo u embrionalnom ili larvalnom razvoju - provizorno. Primjeri privremenih organa su škrge larvi vodozemaca, primarni bubreg i embrionalne membrane viših kralježnjaka (amniote).
U istorijskom razvoju, transformacije organa mogu biti progresivne ili regresivne. U prvom slučaju organi se povećavaju u veličini i postaju složeniji u svojoj strukturi, u drugom se smanjuju i njihova struktura postaje jednostavnija.
Ako dva organizma koja se nalaze na različitim nivoima organizacije imaju organe koji su izgrađeni prema jednom planu, smješteni na istom mjestu i razvijaju se na sličan način iz istih embrionalnih rudimenata, onda to ukazuje na odnos ovih organizama. Takvi organi se nazivaju homologno. Homologni organi često obavljaju istu funkciju (na primjer, srce riba, vodozemaca, gmizavaca i sisara), ali se u procesu evolucije funkcije mogu mijenjati (na primjer, prednji udovi riba i vodozemaca, gmazova i ptica).
Kada nepovezani organizmi žive u istom okruženju, oni mogu razviti slične adaptacije, koje se manifestiraju u izgledu slično organi. Slični organi obavljaju iste funkcije, ali se njihova struktura, lokacija i razvoj oštro razlikuju. Primjeri takvih organa su krila insekata i ptica, udovi i vilični aparat artropoda i kralježnjaka.
Struktura organa striktno odgovara funkcijama koje obavljaju. Štoviše, u povijesnim transformacijama organa, promjena funkcija svakako je praćena promjenom morfoloških karakteristika organa.