Koje su tri vrste membranskih proteina? Integralni i periferni membranski proteini. Pričvršćivanje membranskim "sidrom"
Budućnost medicine su personalizovane metode selektivnog uticaja na pojedinačne ćelijske sisteme koji su odgovorni za razvoj i tok određene bolesti. Glavna klasa terapeutskih meta su proteini stanične membrane kao strukture odgovorne za osiguravanje direktnog prijenosa signala u ćeliju. Već danas gotovo polovina lijekova djeluje specifično na ćelijske membrane, a u budućnosti će ih biti još više. Upoznavanje biološka uloga Ovaj članak je posvećen membranskim proteinima.
Struktura i funkcija stanične membrane
Od školski kurs mnogi se sjećaju uređaja strukturna jedinica organizam - ćelije. Posebno mjesto u strukturi žive ćelije zauzima plazmalema (membrana), koja odvaja unutarćelijski prostor od okoline. Stoga je njegova glavna funkcija stvaranje barijere između ćelijskog sadržaja i ekstracelularnog prostora. Ali to nije jedina funkcija plazmaleme. Ostale membranske funkcije prvenstveno povezane s membranskim proteinima uključuju:
- Zaštitni (vezivanje antigena i sprečavanje njihovog prodora u ćeliju).
- Transport (obezbeđivanje metabolizma između ćelije i okoline).
- Signal (ugrađeni receptorski proteinski kompleksi obezbeđuju razdražljivost ćelije i njen odgovor na različite spoljne uticaje).
- Energija - transformacija različitih oblika energije: mehaničke (bičevi i treplje), električne (nervni impuls) i hemijske (sinteza molekula adenozin trifosforne kiseline).
- Kontakt (omogućavanje komunikacije između stanica pomoću dezmosoma i plazmodezma, kao i nabora i izraslina plazmaleme).

Struktura membrane
Stanična membrana je dvostruki sloj lipida. Dvosloj nastaje zbog prisustva u molekulu lipida dva dijela s različitim svojstvima - hidrofilnog i hidrofobnog područja. Spoljni sloj membrane formiraju polarne „glave” sa hidrofilnim svojstvima, a hidrofobni „repovi” lipida su usmereni u dvosloj. Pored lipida, struktura membrane uključuje proteine. Godine 1972. američki mikrobiolozi S.D. Singer (S. Jonathan Singer) i G.L. Nicholson (Garth L. Nicolson) je predložio fluidno-mozaični model strukture membrane, prema kojem proteini membrane „plutaju“ u lipidnom dvosloju. Ovaj model je proširio njemački biolog Kai Simons (1997) u smislu formiranja određenih, gušćih regija sa pridruženim proteinima (lipidnim splavovima), koji slobodno plivaju u dvosloju membrane.
Prostorna struktura membranskih proteina
IN različite ćelije odnos lipida i proteina je različit (od 25 do 75% proteina u odnosu na suvu masu), a oni su neravnomjerno raspoređeni. Po lokaciji, proteini mogu biti:
- Integralni (transmembranski) - ugrađeni u membranu. Istovremeno, prodiru kroz membranu, ponekad više puta. Njihovi ekstracelularni regioni često nose lance oligosaharida, formirajući glikoproteinske klastere.
- Periferna - nalazi se uglavnom na unutrašnjoj strani membrana. Komunikacija sa membranskim lipidima je osigurana reverzibilnim vodoničnim vezama.
- Usidren - pretežno smješten na vanjskoj strani ćelije i "sidro" koje ih drži na površini je lipidni molekul uronjen u dvosloj.

Funkcionalnost i odgovornosti
Biološka uloga membranskih proteina je raznolika i ovisi o njihovoj strukturi i lokaciji. Među njima su receptorski proteini, kanalni proteini (joni i porini), transporteri, motori i strukturni proteinski klasteri. Svi tipovi proteina membranskih receptora, kao odgovor na bilo koji udar, mijenjaju svoju prostornu strukturu i formiraju ćelijski odgovor. Na primjer, inzulinski receptor regulira protok glukoze u ćeliju, a rodopsin u osjetljivim stanicama organa vida pokreće kaskadu reakcija koje dovode do pojave nervnog impulsa. Uloga proteina membranskih kanala je da transportuju ione i održavaju njihovu razliku između unutrašnjeg i spoljašnjeg okruženja. Na primjer, natrijum-kalijum pumpe osiguravaju razmjenu odgovarajućih jona i tvari. Porini - proteini s kraja na kraj - uključeni su u prijenos molekula vode, transporteri - u prijenos određenih tvari protiv gradijenta koncentracije. Kod bakterija i protozoa, kretanje flagele osiguravaju molekularni proteinski motori. Strukturni membranski proteini podržavaju samu membranu i osiguravaju interakciju drugih proteina plazma membrane.

Proteini za membranu, membrana za proteine
Membrana je dinamično i vrlo aktivno okruženje, a ne inertna matrica za proteine koji se nalaze i rade u njoj. Značajno utječe na funkcioniranje membranskih proteina, a lipidni splavi, krećući se, formiraju nove asocijativne veze proteinskih molekula. Mnogi proteini jednostavno ne rade bez partnera, a njihova intermolekularna interakcija je osigurana prirodom lipidnog sloja membrane, čija strukturna organizacija, zauzvrat, ovisi o strukturnim proteinima. Poremećaji u ovom delikatnom mehanizmu interakcije i međuzavisnosti dovode do disfunkcije membranskih proteina i niza bolesti, poput dijabetesa i malignih tumora.
Strukturna organizacija
Moderne ideje o strukturi i strukturi membranskih proteina zasnivaju se na činjenici da se u perifernom dijelu membrane većina njih rijetko sastoji od jednog, češće od nekoliko povezanih oligomerizirajućih alfa heliksa. Štoviše, upravo je ova struktura ključ za obavljanje funkcije. Međutim, klasifikacija proteina prema vrsti strukture može donijeti još mnogo iznenađenja. Od više od stotinu opisanih proteina, najviše proučavan membranski protein prema vrsti oligomerizacije je glikoforin A (protein crvenih krvnih zrnaca). Kod transmembranskih proteina situacija je složenija – opisan je samo jedan protein (fotosintetski reakcijski centar bakterija – bakteriorhodopsin). S obzirom na visoku molekularnu težinu membranskih proteina (10-240 hiljada daltona), molekularni biolozi imaju široko polje istraživanja.
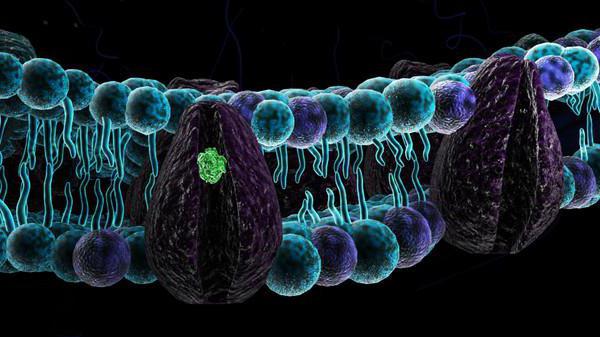
Sistemi ćelijske signalizacije
Među svim proteinima plazma membrane, posebno mjesto pripada receptorskim proteinima. Oni regulišu koji signali ulaze u ćeliju, a koji ne. Kod svih višećelijskih organizama i nekih bakterija informacije se prenose putem posebnih molekula (signalnih molekula). Među ovim signalnim agensima su i oni posebno izlučeni od strane ćelija), neproteinske formacije i pojedinačni joni. Potonje se može osloboditi kada su susjedne ćelije oštećene i pokreću kaskadu reakcija u obliku bola, glavnog odbrambenog mehanizma tijela.
Ciljevi za farmakologiju
Upravo su membranski proteini glavne mete farmakologije, budući da su one tačke kroz koje prolazi većina signala. "Ciljanje" lijeka i osiguranje njegove visoke selektivnosti glavni je zadatak pri stvaranju farmakološkog sredstva. Selektivno djelovanje samo na određeni tip ili čak podtip receptora je djelovanje samo na jednu vrstu ćelije u tijelu. Takvo selektivno djelovanje može, na primjer, razlikovati tumorske ćelije od normalnih ćelija.

Lijekovi budućnosti
Svojstva i karakteristike membranskih proteina se već danas koriste u stvaranju lijekova nove generacije. Ove tehnologije se zasnivaju na stvaranju modularnih farmakoloških struktura od nekoliko molekula ili nanočestica međusobno „povezanih”. Dio „ciljanja“ prepoznaje određene receptorske proteine na ćelijskoj membrani (na primjer, one povezane s razvojem raka). Ovom dijelu se dodaje agens koji razbija membranu ili blokator procesa proizvodnje proteina u ćeliji. Razvoj apoptoze (programa samosmrti) ili drugog mehanizma kaskade intracelularnih transformacija dovodi do željenog rezultata izlaganja farmakološkom agensu. Kao rezultat, imamo lijek sa minimalnim nuspojavama. Prvi takvi lijekovi protiv raka već prolaze klinička ispitivanja i uskoro će postati ključ visoko efikasne terapije.

Strukturna genomika
Moderna nauka o proteinskim molekulima se sve više kreće informacione tehnologije. Opsežan put istraživanja - proučiti i opisati sve što je moguće, pohraniti podatke u kompjuterske baze podataka i potom tražiti načine za primjenu tog znanja - cilj je modernih molekularnih biologa. Prije samo petnaest godina pokrenut je globalni projekat ljudskog genoma i već imamo sekvenciranu mapu ljudskih gena. Drugi projekat, čiji je cilj utvrđivanje prostorne strukture svih "ključnih proteina", strukturne genomike, još je daleko od završetka. Prostorna struktura je do sada utvrđena samo za 60 hiljada od više od pet miliona ljudskih proteina. I iako su naučnici do sada uzgajali samo svijetleće prasad i paradajz otporan na hladnoću sa genom lososa, tehnologije strukturne genomike ostaju faza naučnog saznanja, čija praktična primjena neće dugo trajati.
PREDAVANJE
TEMA: „Uvod u histologiju. Plazma membrana, struktura i funkcije. Strukture koje formira plazma membrana"
Histologija, bukvalno prevedena, je nauka o tkivima, ali ovaj koncept ne prihvata zaista veliku količinu materijala koju ova istinski medicinska disciplina pokriva. Kurs histologije počinje proučavanjem citologije ne toliko na svjetlosno-optičkom, koliko na molekularnom nivou, koji moderne medicine logično ušao u etiologiju i patogenezu niza bolesti. Histologija također uključuje posebne dijelove iz kursa embriologije, ne sve, naravno, nego onaj dio koji se dotiče pitanja formiranja i diferencijacije primordija tkiva. I na kraju, histologija je veliki dio privatne histologije, odnosno dio koji proučava strukturu i funkcije različitih organa. Navedeni dijelovi predmeta histologija ne ostavljaju nikakvu sumnju da izučavanje naše discipline treba izvoditi u aspektu održavanja jedinstva ćelijskog, tkivnog, organskog i sistemskog nivoa organizacije.
Histologiju započinjemo proučavanjem eukariotske ćelije, koja je najjednostavniji sistem obdaren životom. Prilikom ispitivanja ćelije pod svetlosnim mikroskopom, dobijamo informacije o njenoj veličini, obliku, a ta informacija je povezana sa prisustvom granica vezanih za membranu u ćelijama. Sa razvojem elektronske mikroskopije (EM), naše razumevanje membrane kao jasno definisane linije razdvajanja između ćelije i okruženje promijenio, jer se pokazalo da na površini ćelije postoji složena struktura, koji se sastoji od sljedećeg 3 komponente:
1. Supramembranska komponenta (glikokaliks) (5-100 nm)
2. Plazma membrana (8-10 nm)
3. Submembranska komponenta (zona varijacije proteina citoskeleta)
Istovremeno, komponente 1 i 3 su promjenjive i zavise od tipa ćelije; čini se da je najstatičnija struktura plazma membrane, koju ćemo razmotriti.
Proučavanje plazmaleme u EM uslovima dovelo je do zaključka da je njena strukturna organizacija ujednačena, u kojoj ima izgled trilaminarne linije, gde su unutrašnji i spoljašnji slojevi elektronski gusti, a širi sloj koji se nalazi između njih izgleda kao da biti elektron-transparentan. Ova vrsta strukturne organizacije membrane ukazuje na njenu hemijsku heterogenost. Ne dotičući se rasprave o ovom pitanju, odredit ćemo da se plazmalema sastoji od tri vrste tvari: lipida, proteina i ugljikohidrata.
Lipidi, koji su dio membrana, imaju amfifilna svojstva zbog prisustva u njihovom sastavu i hidrofilnih i hidrofobnih grupa.
Amfipatska priroda membranskih lipida potiče stvaranje lipidnog dvosloja. U ovom slučaju razlikuju se dva domena u membranskim fosfolipidima: a) fosfat– glava molekula, Hemijska svojstva ovaj domen određuje njegovu rastvorljivost u vodi i naziva se hidrofilnim
b) acilni lanci, koje su esterifikovane masne kiseline – ovo je hidrofobni domen.
Vrste membranskih lipida. 1. Glavna klasa lipida u biološkim membranama su fosfo(gliceridi) (fosfolipidi), oni čine okvir
biološka membrana (slika 1).
Biomembrane- ovo je dvostruki sloj amfifilnih lipida(lipidni dvosloj). IN vodena sredina takvi amfifilni molekuli spontano formiraju dvosloj u kojem su hidrofobni dijelovi molekula orijentirani jedan prema drugom, a hidrofilni prema vodi (slika 2).
Membrane sadrže sljedeće vrste lipida:
1. Fosfolipidi
2. Sfingolipidi "glava" + 2 hidrofobna "repa"
3.Glikolipidi
kolesterol (CL)– nalazi se u membrani uglavnom u srednjoj zoni dvosloja, amfifilna je i hidrofobna(osim jedne hidroksi grupe). Sastav lipida utiče na svojstva membrana: odnos protein/lipid je blizu 1:1, međutim, mijelinske ovojnice su obogaćene lipidima, a unutrašnje membrane su obogaćene proteinima.
Metode pakovanja amfifilnih lipida: 1. Dvoslojevi (lipidna membrana), 2. Liposomi su vezikule sa dva sloja lipida, dok su i unutrašnja i vanjska površina polarne. 3. Micele - treća varijanta organizacije amfifilnih lipida - vezikula, čiji zid čini jedan sloj lipida, dok su njihovi hidrofobni krajevi okrenuti ka centru micele i njihova unutrašnja sredina nije vodena, već hidrofobna.
Najčešći oblik pakovanja molekula lipida je njihovo formiranje stan membranski dvosloj. Liposomi i micele su brzi transportni oblici koji osiguravaju prijenos tvari u i iz stanice. U medicini se liposomi koriste za transport materija rastvorljivih u vodi, a micele se koriste za transport supstanci rastvorljivih u mastima.
Membranski proteini:
1. Integralni (uključen u slojeve lipida)
2. Periferni
Integralni (transmembranski proteini):
1. Monotop- (npr. glikoforin. 1 put prelaze membranu), i receptori su, dok njihov spoljni - ekstracelularni domen - pripada prepoznavnom delu molekula.
2. Polytopic– više puta prodiru kroz membranu – to su također receptorski proteini, ali aktiviraju put prijenosa signala u ćeliju.
Membranski proteini povezani s lipidima.
4. Membranski proteini, povezana sa ugljikohidratima.
Periferni proteini - nisu uronjeni u lipidni dvosloj i nisu kovalentno vezani za njega. Oni se drže zajedno ionskim interakcijama. Periferni proteini su povezani sa integralnim proteinima u membrani zbog interakcije - protein-protein interakcije.
Primjer ovih proteina:
1. Spektrin, koji se nalazi na unutrašnja površinaćelije
2. fibronektin, lokaliziran na vanjskoj površini membrane
proteini – obično čine do 50% mase membrane. Gde
integralni proteini obavljaju sljedeće funkcije:
a) proteini jonskih kanala
b) receptorske proteine
2. Proteini periferne membrane(fibrilarni, globularni) obavljaju sljedeće funkcije:
a) vanjski (receptorski i adhezioni proteini)
b) unutrašnji – proteini citoskeleta (spektrin, ankirin), proteini drugog glasničkog sistema.
Jonski kanali– to su kanali formirani od integralnih proteina; oni formiraju male pore kroz koje ioni prolaze duž elektrohemijskog gradijenta. Najpoznatiji kanali su kanali za Na, K, Ca 2, Cl.
Postoje i vodeni kanali - to su akvaporini(eritrociti, bubrezi, oko).
Supramembranska komponenta– glikokaliks, debljine 50 nm. To su ugljikohidratni dijelovi glikoproteina i glikolipida koji daju negativan naboj. Ispod EM je labav sloj umjerene gustoće koji pokriva vanjsku površinu plazmaleme. Osim komponenti ugljikohidrata, glikokaliks sadrži proteine periferne membrane (poluintegralne). Njihova funkcionalna područja nalaze se u supramembranskoj zoni - to su imunoglobulini (slika 4).
Funkcija glikokaliksa: 1. Igrajte ulogu receptori.
2. Intercelularno prepoznavanje.
3. Međućelijske interakcije(ahezivne interakcije).
4. R receptore histokompatibilnosti.
5. Zona adsorpcije enzima(parietalna probava).
6. Hormonski receptori.
Submembranska komponenta ili najudaljenija zona citoplazme, obično ima relativnu krutost i ova zona je posebno bogata filamentima (d 5-10 nm). Pretpostavlja se da su integralni proteini koji čine staničnu membranu direktno ili indirektno povezani sa aktinskim filamentima koji leže u submembranskoj zoni. Istovremeno, eksperimentalno je dokazano da se tokom agregacije integralnih proteina aktin i miozin koji se nalaze u ovoj zoni takođe agregiraju, što ukazuje na učešće aktinskih filamenata u regulaciji oblika ćelije.
Strukture formirane plazmalemom
Konture ćelije, čak ni na svjetlosno-optičkom nivou, ne izgledaju ujednačene i glatke, a elektronska mikroskopija je omogućila otkrivanje i opisivanje različitih struktura u ćeliji koje odražavaju prirodu njene funkcionalne specijalizacije. Razlikuju se sljedeće strukture:
1. Microvilli – izbočenje citoplazme prekrivene plazmalemom. Citoskelet mikrovilusa je formiran od snopa aktinskih mikrofilamenata, koji su utkani u terminalnu mrežu apikalnog dijela ćelija (slika 5). Pojedinačne mikroresice nisu vidljive na svjetlosnom optičkom nivou. Ako ih ima značajan broj (do 2000-3000) u apikalnom dijelu ćelije, čak i uz svjetlosnu mikroskopiju razlikuje se "granica četkice".
2. Trepavice – nalaze se u apikalnoj zoni ćelije i imaju dva dela (slika 6): a) spoljašnji - aksonem
B) unutrašnje - bekalno tijelo
Axoneme sastoji se od kompleksa mikrotubula (9 + 1 par) i povezanih proteina. Mikrotubule formira protein tubulin, a ručke formira protein dinein - ovi proteini zajedno formiraju tubulin-dinein hemomehanički pretvarač.
Bazalno tijelo sastoji se od 9 trojki mikrotubula smještenih u bazi cilije i služi kao matrica za organiziranje aksonema.
3. Bazalni labirint- To su duboke invaginacije bazalne plazmaleme sa mitohondrijama koje se nalaze između njih. Ovo je mehanizam za aktivnu apsorpciju vode, kao i jona protiv gradijenta koncentracije.
1. Transport jedinjenja male molekularne težine izvodi se na tri načina:
1. Jednostavna difuzija
2. Olakšana difuzija
3. Aktivni transport
Jednostavna difuzija– hidrofobna organska jedinjenja male molekulske mase (masne kiseline, urea) i neutralni molekuli (HO, CO, O). Kako se povećava razlika u koncentraciji između odjeljaka razdvojenih membranom, tako se povećava i brzina difuzije.
Olakšana difuzija– tvar prolazi kroz membranu također u smjeru gradijenta koncentracije, ali uz pomoć transportnog proteina – translocases. To su integralni proteini koji imaju specifičnost za transportirane supstance. To su, na primjer, anjonski kanali (eritrocit), K kanali (plazmolema ekscitiranih ćelija) i Ca kanali (sarkoplazmatski retikulum). Translocase za H O je akvaporin.
Mehanizam djelovanja translokaze:
1. Prisustvo otvorenog hidrofilnog kanala za supstance određene veličine i naboja.
2. Kanal se otvara samo kada se veže određeni ligand.
3. Kanal kao takav ne postoji, a sam molekul translokaze, nakon što je vezao ligand, rotira za 180 u ravni membrane.
Aktivan transport– ovo je transport koristeći isti transportni protein (translokacije), ali protiv gradijenta koncentracije. Za ovaj pokret je potrebna energija.
Za razliku od lipida, membranske proteine je teško klasificirati po strukturi; prikladnije je podijeliti glavne tipove membranskih proteina na osnovu njihovih funkcija. U pravilu su proteini odgovorni za funkcionalnu aktivnost membrana. Ovi proteini uključuju razne enzime, transportne proteine, receptore, proteine kanala, proteine koji formiraju vodene pore (akvaporine), kao i različite strukturne i regulatorne proteine koji obezbeđuju različite funkcije ćelijskih membrana. Na osnovu njihove biološke uloge, membranski proteini se mogu podijeliti u četiri grupe:
1) enzimski proteini sa katalitičkom aktivnošću;
2) receptorski proteini koji specifično vezuju određene supstance;
3) strukturni proteini;
4) proteini koji osiguravaju međućelijske interakcije.
Enzimski proteini najzastupljeniji od svih membranskih proteina. To uključuje i integralne (membranske ATPaze koje obavljaju transportnu funkciju) i periferne (acetilholinesteraza, kisele i alkalne fosfataze, RNKaze) proteine. Transportni proteini stvaraju stabilne tokove određene supstance i joni kroz membrane. Transport jona dovodi do pojave transmembranskog potencijala u svim ćelijama, kao i do njegovih promena u nervnim i mišićnim ćelijama. Potonji fenomen leži u osnovi tako važnih svojstava ovih tkiva kao što su ekscitabilnost i provodljivost.
Receptorski proteini su proteini koji specifično vezuju određene ligande uključene u prijenos signala iz jedne ćelije u drugu.Takav prijenos se odvija na različite načine. Receptori su često dio složenijih membranskih kompleksa koji sadrže proteine dželata. Na primjer, u nervnim i neuromuskularnim sinapsama signalna molekula (medijator) je određena niskomolekularna supstanca, a plazma membrana sadrži posebne receptorske proteine povezane s ionskim kanalima koji mijenjaju svoja svojstva kada se receptor veže za ligande. Ova reakcija osigurava propusnost membrane za različite ione (natrijum, kalij, kalcij, hlor) i formira uzbudljiv potencijal. Neki receptori (na primjer, nikotinski holinergički receptor) su sami po sebi jonski kanali (zbog uključivanja dodatnih proteinskih podjedinica u receptorski ansambl).
U nekim slučajevima, protein receptora nije jonski kanal, već je povezan sa intracelularnom signalnom kaskadom, do čije aktivacije dolazi nakon vezivanja liganda koji nosi informaciju za receptor. Kao rezultat aktivacije takvih receptora (oni se nazivaju metabotropni, za razliku od jonotropnih, koji reguliraju protok jona kroz membranu), dolazi do kaskade hemijske reakcije, kontroliranje ćelijskih funkcija kroz promjene u metabolizmu (otuda naziv ovih receptora). Aktivacija metabotropnih receptora ligandima (oni se mogu smatrati primarnim signalnim molekulima, ili primarnim glasnicima) dovodi do proizvodnje sekundarnih signalnih molekula (drugih glasnika) u citoplazmi aktivirane ćelije.
Strukturni proteini daju ćelijama i organelama njihov specifičan oblik; pružiti jedno ili drugo mehanička svojstva(npr. elastičnost) plazma membrana; Oni povezuju membranu sa citoskeletom, au slučaju nuklearne membrane sa hromozomima. Proteini strukturne membrane, u pravilu, nemaju enzimska svojstva (možda su jednostavno slabo proučeni kemijski). Njihovo proučavanje ometaju uglavnom dvije okolnosti. Prvo, strukturni proteini su "tihi" - nemaju poznatu enzimsku aktivnost. Drugo, strukturni proteini imaju ekstenzivne hidrofobne regije unutar svojih molekula. Kada se pročiste, formiraju bliske saradnike međusobno ili sa lipidima, što otežava njihovo proučavanje.
Neurospecifični protein B-50 jedan je od glavnih fosforiliranih strukturnih proteina plazma membrana sinaptičkih kontakata. Imunohemijskim metodama utvrđeno je da je lokaliziran pretežno u presinaptičkim membranama. Molekularna masa proteina 48 kPa. To je endogeni supstrat protein kinaze C zavisne protein kinaze C. Aktivatori protein kinaze C stimulišu proces sinaptičke transmisije u hipokampalnim kriškama. Fosforilacija proteina B-50 dovodi do povećanja vremena pobuđenog stanja sinapse, što pomaže u održavanju jonskih kanala u aktiviranom (otvorenom) stanju (u nekim publikacijama ovaj fenomen se naziva stanje sinapse). Utjecaj fosforiliranog proteina B-50 na metabolizam fosfoinozitida može biti jedan od razloga za ovaj fenomen. Zanimljivo je da kako tijelo stari, intenzitet fosforilacije proteina B-50 u mozgu opada, što može uzrokovati smanjenje plastičnosti sinapsi.
Daljnji dokazi o ulozi fosforilacije proteina B-50 u funkcionisanju sinapsi dobijeni su u in vitro eksperimentima, koji su potvrdili da je neuropeptid - fragment ACTH1_24 - 10 puta efikasniji u inhibiciji fosforilacije B-50 u sinaptičke membrane iz septalne regije mozga nego u membranama cijelog mozga
Grupa membranskih proteina uključuje i mnoge enzimske proteine koji formiraju jonske kanale - Na/K- i Ca-ATPaze, receptorske proteine, sinapsine itd.
Plasmin- serin proteinaza, u krvnoj plazmi djeluje uglavnom kao trombolitički enzim, a također razgrađuje mnoge komponente ekstracelularnog matriksa. U mozgu, plazmin je uključen u brojne funkcije kao što su plastičnost neurona, učenje i pamćenje. Aktivacija plazminogenog sistema se opaža u mozgu tokom i prvih dana nakon moždanog udara. Kod Alchajmerove bolesti, naprotiv, dolazi do smanjenja nivoa plazmina u moždanom tkivu.
Plazmin, koji je dio plazma membrane neurona, povezan je sa splavovima bogatim holesterolom, za koje se smatra da su mjesto dominantnog stvaranja amiloida beta-A. Ovo ukazuje na funkcionalnu vezu između plazmina, holesterola i metabolizma mozga.
Enzim koji pretvara endotepin(ECE-1) je još jedan enzim koji razgrađuje amiloid, koji je 37% homologan NEP-u u sekvenci aminokiselina.ECE-1 je također membranski vezana metaloproteaza zavisna od cinka, baš kao i NEP, sposobna je razgraditi veliki broj biološki aktivne supstance, uključujući bradikinin, neurotenzin, angiotenzin-1 i lanac inzulina B. Za razliku od NEP-a, ECE-1 postoji u obliku dimera, čije su podjedinice povezane disulfidnom vezom.
Ove dvije metaloproteaze (ECE-1 i NEP) se također razlikuju po osjetljivosti na inhibitore. Za inhibiciju NEP-a potrebne su nanomolarne koncentracije tiorfana i fosforamidona, dok je ECE-1 inhibiran mikromolarnim koncentracijama fosforamidona i nije osjetljiv na tiorfan.
ECE-1 se nalazi u mnogim organima i tkivima. Endotelne ćelije su najviše obogaćene ovim enzimom, on je takođe izražen u nervnog tkiva i mišiće. U ćelijama glatkih mišića, ECE-1 se nalazi u kompleksu sa alfa-aktinskim filamentima.
Trenutno su poznate četiri ljudske ECE-1 izofore (1a, 1b, 1c i 1d), koje nemaju značajne katalitičke razlike, ali se razlikuju u subćelijskoj lokalizaciji. Izoforme 1a, 1b, 1c i 1d nalaze se na površini ćelije, dok je ECE-1 intracelularni oblik lokalizovan u Golgijevom aparatu.
Otkriven je još jedan protein sličan ECE-1, koji je lokaliziran uglavnom u mozgu iu malim količinama u endotelnim i glatkim mišićnim stanicama. Njegova primarna struktura je 59°/o identična sekvenci aminokiselina ECE-1. Označen je kao ECE-2 i razlikuje se od ECE-1 po tome što ima kiseliji pH optimum.
| | | | | | | | | 10 |
U pravilu su proteini odgovorni za funkcionalnu aktivnost membrana. To uključuje razne enzime, transportne proteine, receptore, kanale, pore, itd. Ranije se vjerovalo da membranski proteini imaju isključivo β presavijenu sekundarnu strukturu proteina, ali ove studije su pokazale da membrane sadrže veliki broj α spirala. Dalja istraživanja su pokazala da membranski proteini mogu prodrijeti duboko u lipidni dvosloj ili čak prodrijeti u njega, a njihova stabilizacija se vrši zahvaljujući hidrofobnom...
Podijelite svoj rad na društvenim mrežama
Ako vam ovaj rad ne odgovara, na dnu stranice nalazi se lista sličnih radova. Možete koristiti i dugme za pretragu
Predavanje 5
Struktura i funkcije membranskih proteina
Ćelijske membrane sadrže proteine od 20 do 80% (po težini). U pravilu su proteini odgovorni za funkcionalnu aktivnost membrana. To uključuje razne enzime, transportne proteine, receptore, kanale, pore, itd. itd., koji osiguravaju jedinstvene funkcije svake membrane. Prvi napredak u proučavanju membranskih proteina postignut je kada su biohemičari naučili koristiti deterdžente za izolaciju membranskih proteina u funkcionalno aktivnom obliku. Radilo se o radovima na proučavanju enzimskih kompleksa unutrašnje membrane mitohondrija. Ranije se vjerovalo da membranski proteini imaju isključivo β presavijenu strukturu (proteinska sekundarna struktura), ali ove studije su pokazale da membrane sadrže veliki broj α spirala. β spirala je mnogo rjeđa, ali joj se, međutim, pridaje važno biološki značaj. Činjenica je da je u područjima okruženim lipidima β spirala šuplji cilindar, u čijem su vanjskom zidu koncentrirani nepolarni (hidrofobni) ostaci aminokiselina, a u unutrašnjim hidrofilni. Takav cilindar mogao bi formirati kanal u membrani kroz koji slobodno prolaze ioni i tvari rastvorljive u vodi. Dalja istraživanja su pokazala da membranski proteini mogu prodrijeti duboko u lipidni dvosloj ili čak prodrijeti u dvosloj lipida i stabiliziraju se hidrofobnim interakcijama. Postoje najmanje četiri tipa rasporeda proteina u membranama: Prvi tip je transmembranski, kada protein prodire kroz cijelu membranu, a hidrofobna regija proteina ima α konfiguraciju. Molekul bakteriorhodopsina iz Halobacterium halobium njegove α spirale sekvencijalno prelaze dvosloj; Drugi tip je vezivanje pomoću hidrofobnog sidra, kada protein ima kratku regiju koja se sastoji od hidrofobnih aminokiselinskih ostataka blizu karboksilnog kraja. Ovo je takozvano hidrofobno sidro, koje se može ukloniti proteolizom, a oslobođeni protein postaje topiv u vodi. Ovaj raspored u membrani karakterističan je za mnoge citohrome. Treći tip je vezivanje za površinu dvosloja, kada je interakcija proteina prvenstveno elektrostatičke ili hidrofobne prirode. Ova vrsta interakcije može se koristiti kao dodatak drugim interakcijama, kao što je transmembransko sidrenje. Četvrti tip je vezivanje za proteine ugrađene u dvosloj, a to je kada se neki proteini vežu za proteine koji se nalaze unutar lipidnog dvosloja. Na primjer, F 1 - dio H + - ATPaza koja se vezuje za F 0 dio uronjen u membranu, kao i neki proteini citoskeleta.
U srži moderne ideje Struktura membranskih proteina zasniva se na ideji da je njihov polipeptidni lanac presavijen tako da formira nepolarnu, hidrofobnu površinu u kontaktu sa nepolarnim regionom lipidnog dvosloja. Polarni domeni proteinske molekule mogu stupiti u interakciju s polarnim glavama lipida na površini dvosloja. Mnogi proteini su transmembranski i obuhvataju dvosloj. Čini se da su neki proteini povezani s membranom samo kroz njihovu interakciju s drugim proteinima.
Mnogi membranski proteini se obično povezuju s membranom kroz nekovalentne interakcije. Međutim, postoje proteini koji su kovalentno vezani za lipide. Mnogi proteini plazma membrane pripadaju klasi glikoproteina. Ostaci ugljikohidrata ovih proteina uvijek se nalaze na vanjskoj strani plazma membrane.
Tipično, membranski proteini se dijele na vanjske (periferne) i unutrašnje (integralne). U ovom slučaju, kriterijum je stepen ozbiljnosti obrade neophodne za ekstrakciju ovih proteina iz membrane. Periferni proteini se oslobađaju kada se membrane isperu puferskim rastvorima niske jonske snage, niskog ili, obrnuto, visokog pH i u prisustvu helatnih agenasa (na primer, EDTA) koji vežu dvovalentne katione. Često je vrlo teško razlikovati proteine periferne membrane od proteina vezanih za membranu tokom procesa oslobađanja.
Za oslobađanje integralnih membranskih proteina potrebno je koristiti deterdžente ili čak organska otapala.
Mnogi proteini eukariotske i prokariotske membrane su kovalentno vezani za lipide, koji se dodaju polipeptidu nakon translacije.
Membranski proteini kovalentno povezani sa lipidima
(A) Proteini za koje je vezana miristinska kiselina
(B) Proteini za koje je vezana palmitinska kiselina
(IN) Proteini sa sidrom za glikozilfosfatidilinozitol
4. Molekul adhezije nervnih ćelija |
U nekim slučajevima ovi lipidi igraju ulogu hidrofobnog sidra, uz pomoć kojih se protein vezuje za membranu. U drugim slučajevima, lipidi će vjerovatno djelovati kao pomoć u migraciji proteina u odgovarajuću ćelijsku regiju ili (kao u slučaju proteina virusne ovojnice) u fuziji membrane.
Kod prokariota, najpotpunije karakterizirani protein je Brownov lipoprotein, glavni lipoprotein vanjske membrane. E. coli . Zreli oblik ovog proteina sadrži acilglicerol, koji je vezan tioeterskom vezom za N terminalni cistein. osim toga, N Terminalna aminokiselina je povezana sa masnom kiselinom amidnom vezom. Za membranu vezan oblik penicilaze je vezan za citoplazmatsku membranu pomoću N terminalni acilglicerol sličan membranskim lipoproteinima.
Membranski proteini eukariota su kovalentno povezani s lipidima, kao što je prikazano u tabeli, mogu se podijeliti u tri klase. Čini se da su proteini prve dvije klase uglavnom lokalizirani na citoplazmatskoj površini plazma membrane, a proteini treće klase na vanjskoj površini.
Postoje membranski proteini koji su kovalentno povezani s ugljikohidratima. To uključuje proteine ćelijske površine koji uglavnom obavljaju funkcije transporta i prijema. Još uvek je nejasno šta se ovde dešava. Ovo može biti zbog činjenice da proteine treba sortirati kada su usmjereni prema plazma membrani. Ostaci šećera mogu zaštititi protein od proteolize ili sudjelovati u prepoznavanju ili adheziji. Stoga su ostaci šećera u membranskim glikoproteinima lokalizirani isključivo na vanjskoj strani membrane.
Mogu se razlikovati dvije glavne klase oligosaharidnih struktura membranskih glikoproteina: 1) N glikozidni oligosaharidi povezani sa proteinima preko amidne grupe aspargina; 2) O-glikozidni oligosaharidi povezani preko hidroksilne grupe serin i treonin. Ova klasa oligosaharida sastoji se od tri podklase.
- Jednostavan kompleks ili kompleks bogat manozom u kojem oligosaharid sadrži manozu i N acetilglukozamin.
- Normalan kompleks u kojem jezgro bogato manozom ima dodatne bočne grane koje sadrže druge ostatke saharida, kao što je sijalinska kiselina.
- Veliki kompleks koji je povezan sa anjonskim transporterom membrane eritrocita
Većina membranskih glikoproteinskih oligosaharida pripada potklasi 1 ili 2.
Membranski proteini bakterija
Kao što je gore navedeno, proteini u citoplazmatskoj membrani čine oko 50% njene površine. Otprilike 10% membrane je formirano od čvrsto vezanih proteinsko-lipidnih kompleksa. Molekul bilo kojeg proteina ugrađenog u membranu okružen je sa 45 × 130 ili više molekula lipida. Otprilike polovina slobodnih lipida je povezana s perifernim membranskim proteinima.
Proteinski sastav citoplazmatske membrane bakterija je raznovrsniji od sastava lipida. Dakle, u citoplazmatskoj membrani E. coli K 12 Otkriveno je oko 120 različitih proteina. Ovisno o orijentaciji u membrani i prirodi veze sa lipidnim dvoslojem, kao što je gore navedeno, proteini se dijele na integralne i periferne. Periferni bakterijski proteini uključuju brojne enzime kao što su NADH dehidrogenaza, malat dehidrogenaza, itd., kao i neke proteine koji su dio kompleksa ATPaze. Ovaj kompleks je grupa proteinskih podjedinica raspoređenih na određeni način, u kontaktu sa citoplazmom, periplazmatskim prostorom i formirajući kanal u membrani kroz koji prolazi proton. Označena površina kompleksa F 1 , uronjen u citoplazmu, i sa komponentama mjesta F 0 Hidrofobne strane molekula su uronjene u membranu. Podjedinica b djelomično uronjen u membranu svojim hidrofobnim dijelom i komunicira membranu i citoplazmatski dio enzimskog kompleksa, kao i vezu sinteze ATP-a u regiji F 1 sa protonskim potencijalom u membrani. podjedinice a, b i c obezbeđuju protonski kanal. Ostale komponente kompleksa osiguravaju njegovu strukturnu i funkcionalnu cjelovitost.
Ka integralnim proteinima E. coli, kojima su potrebni lipidi za ispoljavanje enzimske aktivnosti, uključuju sukcinat dehidrogenazu, citokrom b . Vrlo zanimljiva svojstva imaju antibiotici gramicidin A, alameticin, amfotericin i nistacin. Kada stupe u interakciju s bakterijskom membranom, postaju integralni proteini (antibiotici su polipeptidi i makrocikli).
Gramicidin A je hidrofobni peptid koji se sastoji od 15 L-D -amino kiseline. Kada je ugrađen u membranu, formira kanale koji omogućavaju prolaz monovalentnim kationima. Ovaj kanal, koji formira gramicidin A, je najpotpunije okarakterisan. Kanal formiraju dva molekula gramicidina A. Kao rezultat alternacije L- i D - aminokiseline formiraju spiralu u kojoj su bočni lanci smješteni izvana, a karboksilne grupe kičme unutar kanala. Ova vrsta heliksa nije pronađena ni u jednom drugom proteinu i formirana je od neobične alternacije stereoizomera aminokiselina u gramicidinu A. Gramicidinski kanal, kao što je gore navedeno, je kationski selektivan. Kroz njega prolaze mali neorganski i organski katjoni, a istovremeno prolaze kroz njega i propusnost Cl - jednak je nuli.
Alameticin je peptidni antibiotik od 20 aminokiselinskih ostataka, sposoban za formiranje električnih podražljivih kanala u membrani. Aminokiselinska sekvenca alameticina uključuje neobične ostatke α aminobuterne kiseline i L fenilalanin. Kada se veže za membranu, za razliku od gramicidina A, formira pore. Mnogo je manje veličine od kanala koji formira gramicidin A. To je prvenstveno zbog činjenice da je prostor oko α heliksa premali da bi ion mogao proći.
Markolidni antibiotici kao što su nistatin i amfotericin vezuju se za holesterol i formiraju kanale. Kanali formiraju 8 × 10 molekula ovih polienskih antibiotika, kroz koje, međutim, ioni prodiru malim brzinama.
Ostali slični radovi koji bi vas mogli zanimati.vshm> |
|||
| 21572. | STRUKTURA I FUNKCIJA PROTEINA | 227,74 KB | |
| Sadržaj proteina u ljudskom tijelu veći je od sadržaja lipida u ugljikohidratima. Prevlast proteina u tkivima u odnosu na druge supstance otkriva se pri izračunavanju sadržaja proteina po suvoj masi tkiva. Sadržaj proteina u različitim tkivima varira unutar određenog raspona. | |||
| 17723. | Mali mozak, struktura i funkcije | 22,22 KB | |
| 3 Opća struktura mozga. IN nervni sistem Razlikuju i centralni dio centralnog nervnog sistema, koji je predstavljen mozgom i kičmenom moždinom, i periferni dio, koji uključuje živce, nervne ćelije, nervne čvorove, ganglije i pleksuse, koji se topografski nalaze izvan kičmene moždine i mozga. . Predmet proučavanja je anatomija mozga. Ovaj cilj subjekta i objekta podrazumeva formulisanje i rešavanje sledećih zadataka: opisati opšti plan struktura mozga; proučavanje anatomske strukture malog mozga; isticanje... | |||
| 5955. | Biljni organi: njihove funkcije, struktura i metamorfoze. | 16,94 KB | |
| Cvjetni organi su modificirani listovi: pokrovni listovi formiraju sepale i latice, a listovi koji tvore spore stvaraju prašnike i tučke. Izdanak uključuje: a stabljiku b listove c vegetativne pupoljke d cvjetove e plodove. Stabljika je vegetativni organ biljke koji obavlja brojne funkcije: nosi lišće ili tešku krunu grana i listova; veže korijenje i lišće; na njemu se formiraju cvjetovi; po njoj se kreće voda s mineralima i organskim spojevima; mlade stabljike... | |||
| 5067. | Glatki mišići. Struktura, funkcije, mehanizam kontrakcije | 134,79 KB | |
| Mišići ili mišići iz lat. Mišići vam omogućavaju da pomerate delove tela i izražavate misli i osećanja u delima. Glatki mišići su dio nekih unutrašnje organe i učestvuju u obezbjeđivanju funkcija koje obavljaju ova tijela. | |||
| 6233. | Struktura i funkcije jezgra. Morfologija i hemijski sastav jezgra | 10,22 KB | |
| Jezgra su obično odvojena od citoplazme jasnom granicom. Bakterije i modrozelene alge nemaju formirano jezgro: njihovo jezgro nema nukleolus i nije odvojeno od citoplazme jasno definiranom nuklearnom membranom i naziva se nukleoid. Oblik jezgra. | |||
| 9495. | Klasifikacija, karakteristike asortimana krznenih sirovina i krznenih poluproizvoda, struktura kože krzna, struktura dlake i raznolikost njenih oblika, tehnologija proizvodnje krzna | 1.05 MB | |
| Krznene ploče su trake određenog oblika, sašivene od odabranih obrađenih koža i namijenjene za rezanje na dijelove krznenih proizvoda. Zimske vrste krznenih sirovina uključuju kože i krzna životinja koje nose krzno, čije se vađenje uglavnom vrši u zimsko vrijeme kada je kvalitet kože posebno visok. STRUKTURA I HEMIJSKI SASTAV KRZNA I ŠEPENOVO KRZNO KOŽE SIROVINE POJAM TOPOGRAFIJE KOŽE Koža je spoljašnji omotač životinje, odvojen od trupa i sastoji se od kožnog tkiva i dlake. U... | |||
| 8011. | Osobine membranskih lipida | 10,13 KB | |
| Neki lipidi pomažu u stabilizaciji visoko zakrivljenih područja membrane, stvaraju kontakt između membrana ili vežu određene proteine, budući da oblik ovih molekula pogoduje željenom pakovanju dvosloja u odgovarajućim područjima membrane. Tečno stanje se podrazumijeva kao sposobnost fosfolipidnih molekula da se rotiraju i bočno kreću u odgovarajućem membranskom režnju. Izdužene su i orijentirane okomito na ravan membrane. U tečnom kristalnom stanju, molekuli masnih kiselina su pokretni, ali... | |||
| 8014. | Hemijski sastav membranskih lipida | 10,81 KB | |
| Prije svega, to je zbog mnogih funkcija koje lipidi obavljaju u membranama. Fosfatidna kiselina se nalazi u slobodnom obliku u bakterijskim membranama u malim količinama, obično sa ostacima aminokiselinskih alkohola i drugih vezanih za nju.Ovi lipidi su estri masnih kiselina i glicerola i široko su zastupljeni u mnogim membranama eukariotskih i prokariotskih ćelija sa izuzetak arhebakterija. Nalaze se u velikim količinama u unutrašnjoj membrani mitohondrija, u membrani hloroplasta i u nekim bakterijskim... | |||
| 21479. | METABOLIZAM PROTEINA | 150,03 KB | |
| Postoje tri tipa bilansa azota: bilans azota pozitivan balans azota negativan balans azota Sa pozitivnim balansom azota, unos azota prevladava nad njegovim otpuštanjem. Kod bolesti bubrega moguća je lažno pozitivna ravnoteža dušika, u kojoj se krajnji produkti metabolizma dušika zadržavaju u tijelu. Sa negativnom ravnotežom dušika, izlučivanje dušika prevladava nad njegovim unosom. Ovo stanje je moguće kod bolesti poput tuberkuloze, reume, onkoloških... | |||
| 15073. | Razmatranje membranskih (ion-selektivnih) elektroda sa različitim tipovima membrana | 127,48 KB | |
| U tu svrhu postoje različite ionsko-selektivne elektrode, čija je glavna karakteristika tzv. selektivnost prema određenoj vrsti jona. Elektrode sa tečnim i filmskim membranama Tečne membrane su rastvori u organskim rastvaračima jonoizmenjivačkih supstanci, tečnih kationskih izmenjivača ili anionskih izmenjivača ili neutralnih helata, odvojeni od vodenih rastvora neutralnim poroznim pregradama, polimer staklom ili dr. Trenutno industrija proizvodi film ion- selektivne elektrode za katjone N K NH4 Ca2.. . | |||
Biološka hemija Lelevič Vladimir Valerijanovič
Membranski proteini.
Membranski proteini.
Membranski proteini su odgovorni za funkcionalnu aktivnost membrana i čine 30 do 70%. Membranski proteini se razlikuju po svom položaju u membrani. Mogu prodrijeti duboko u lipidni dvosloj ili čak prodrijeti u njega - integralni proteini, Različiti putevi vežu se za membranu - površinski proteini, ili kovalentno kontaktiraju s njom - usidreni proteini. Površinski proteini su gotovo uvijek glikozilirani. Ostaci oligosaharida štite protein od proteolize i uključeni su u prepoznavanje i adheziju liganda.
Proteini lokalizirani u membrani obavljaju strukturne i specifične funkcije:
1. transport;
2. enzimski;
3. receptor;
4. antigenski.
Iz knjige Napravi bilo koga, ali NE KROKODILA! autor Orsag MihaiŠta je sa vjevericama? Šezdesetih sam više puta pokušavao da imam vjeverice u kući, ali svaki takav pokušaj završavao se na najtužniji način. Nakon nekog vremena vjeverice su oslabile, zadnji udovi su im oduzeti, a nesretne životinje su umrle u grčevima. Isprva sam
Iz knjige Ljudski genom: enciklopedija napisana u četiri slova autor Iz knjige Ljudski genom [Enciklopedija napisana u četiri slova] autor Tarantul Vjačeslav ZalmanovichNe kodiraju svi geni proteine Prije svega, treba napomenuti da pored gena koji kodiraju proteine, genom sadrži i gene koji sintetiziraju RNK, koji nisu mRNA (odnosno, ne kodiraju proteine), ali obavljaju niz nezavisnih važne funkcije u ćelijama. IN
Iz knjige Živi sat od Ward Ritchie11. Vjeverice u kotaču Da bismo se upoznali sa modernim traganjem za rješenjem problema živih satova, okrenimo se istraživanju koje su proveli biolozi u poslednjih godina. Među prvima je, možda, rad Patricije de Courcy. Godine 1955. de Courcy dobija diplomu
Iz knjige Biologija [ Kompletan vodič pripremiti se za Jedinstveni državni ispit] autor Lerner Georgij Isaakovič Iz knjige Krv: Rijeka života [Od drevnih legendi do naučnim otkrićima] autor Isaac AsimovPoglavlje 11 Ovi mobilni proteini Na početku prethodnog poglavlja spomenuo sam da su organske komponente hrane podijeljene u tri grupe. Govorio sam o jednoj od ovih grupa: ugljeni hidrati. Zatim bi bilo logično da pređemo na proteine, jer se njihov metabolizam u organizmu odvija paralelno sa
Iz knjige Logika slučajnosti [O prirodi i porijeklu biološke evolucije] autor Kunin Evgenij ViktorovičPoglavlje 12 Poreklo života. Pojava translacije, replikacije, metabolizma i membrana: biološki, geohemijski i kosmološki pristupi Trans. A. Neizvestny U prethodnom poglavlju raspravljali smo o mogućim scenarijima za pojavu ćelija i (nadajmo se) postigli
Iz knjige Geni i razvoj tijela autor Nejfah Aleksandar Aleksandrovič2. Proteini hromatina Već znamo da se hromatin sastoji od DNK i histona u jednakim količinama i nehistonskih proteina (NGP), kojih u neaktivnim regionima hromozoma ima samo 0,2 težine DNK, au aktivnim regionima - više od 1,2 (u prosjeku NGB je manji od DNK). Takođe znamo da histoni
Iz knjige Životinjski svijet autor Sitnikov Vitalij Pavlovič Iz knjige Trenutna drzava biosfera i ekološka politika autor Kolesnik Yu. A.4.1. Formiranje membrana je osnova za početak života.S obzirom na gigantsku raznolikost modernih živih organizama, možemo doći do zaključka da je postojalo mnogo razvojnih puteva koji potiču od reliktnih oblika života. Zapravo, proučavanje molekularne evolucije
Iz knjige Tajne ljudske nasljednosti autor Afonkin Sergej JurijevičĆelije, proteini i geni Život je način postojanja proteinskih tijela. F. Engels Naše tijelo je carstvo ćelija, od kojih je svaka minijaturna fabrika za proizvodnju proteina. Mnoge od ovih esencijalnih makromolekula mogu se izlučiti iz tijela
Iz knjige Antropologija i koncepti biologije autor Kurčanov Nikolaj AnatolijevičProteini Proteini su od najveće važnosti u životu organizama. Ogromna raznolikost živih bića je u velikoj mjeri određena razlikama u sastavu proteina prisutnih u njihovim tijelima. Na primjer, u ljudskom tijelu ih je poznato više od 5 miliona.Proteini su polimeri,
Iz knjige Biološka hemija autor Lelevič Vladimir ValerijanovičHemijski sastav membrane Membrane se sastoje od molekula lipida i proteina, čije relativne količine uvelike variraju među različitim membranama. Ugljikohidrati se nalaze u obliku glikoproteina, glikolipida i čine 0,5%-10% membranskih supstanci. Prema tečnom mozaiku
Iz knjige autoraMembranski lipidi. Membranski lipidi su amfifilne molekule, tj. molekul sadrži i hidrofilne grupe (polarne glave) i alifatske radikale (hidrofobni repovi), koji spontano formiraju dvosloj u kojem su repovi lipida okrenuti jedan prema drugom. Debljina
Iz knjige autoraProteini Nutritivnu vrijednost bjelančevina osigurava prisustvo esencijalnih aminokiselina, čiji se ugljikovodični kosturi ne mogu sintetizirati u ljudskom tijelu, te se shodno tome moraju snabdjeti hranom. Oni su takođe glavni izvori azota. Dnevnice
Iz knjige autoraProteini mišićnog tkiva Postoje tri grupe proteina: 1. miofibrilarni proteini – 45%;2. sarkoplazmatski proteini – 35%;3. stromalni proteini – 20%.Miofibrilarni proteini.Ova grupa uključuje: 1. miozin; 2. aktin;3. aktomiozin; kao i takozvani regulatorni proteini: 4. tropomiozin;5.





